

Nucleotide-induced switch in oligomerization of the AAA+ ATPase ClpB
- PMID: 14978298
- PMCID: PMC1828688
- DOI: 10.1110/ps.03422604
Nucleotide-induced switch in oligomerization of the AAA+ ATPase ClpB
Abstract
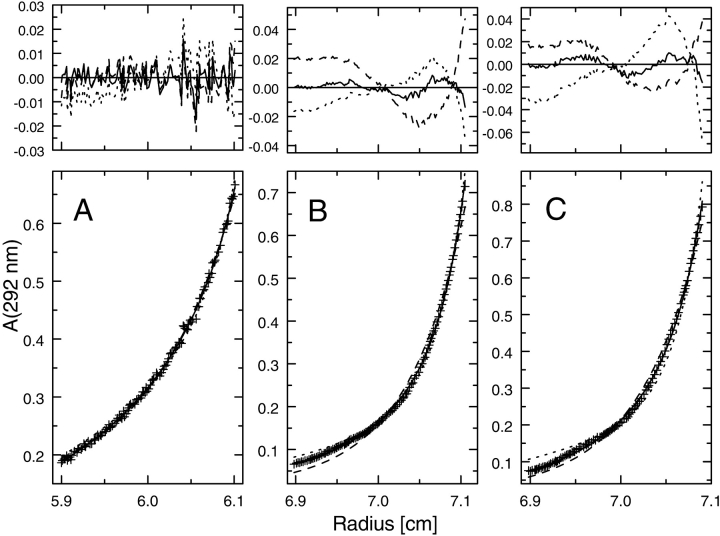
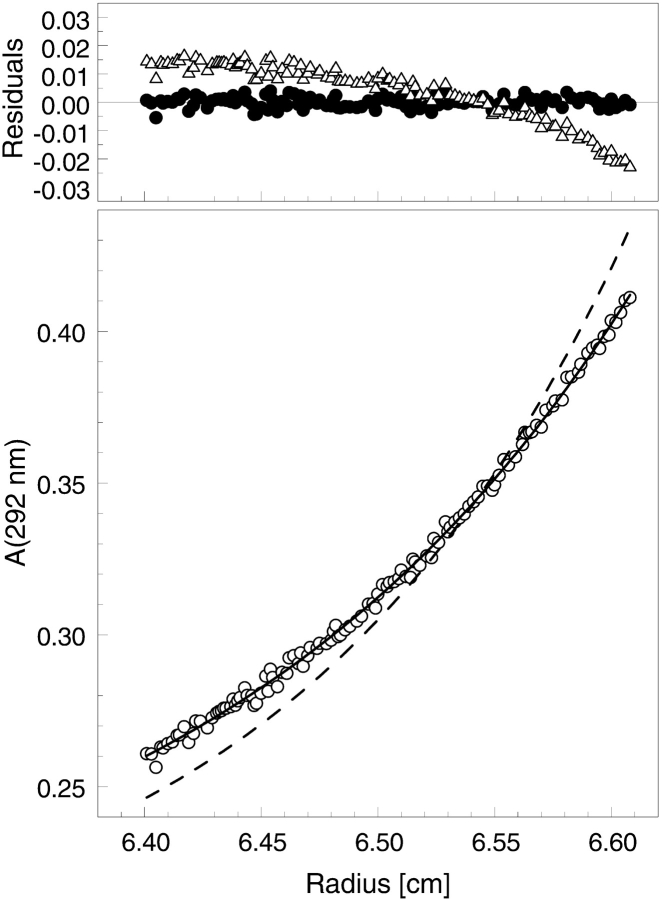
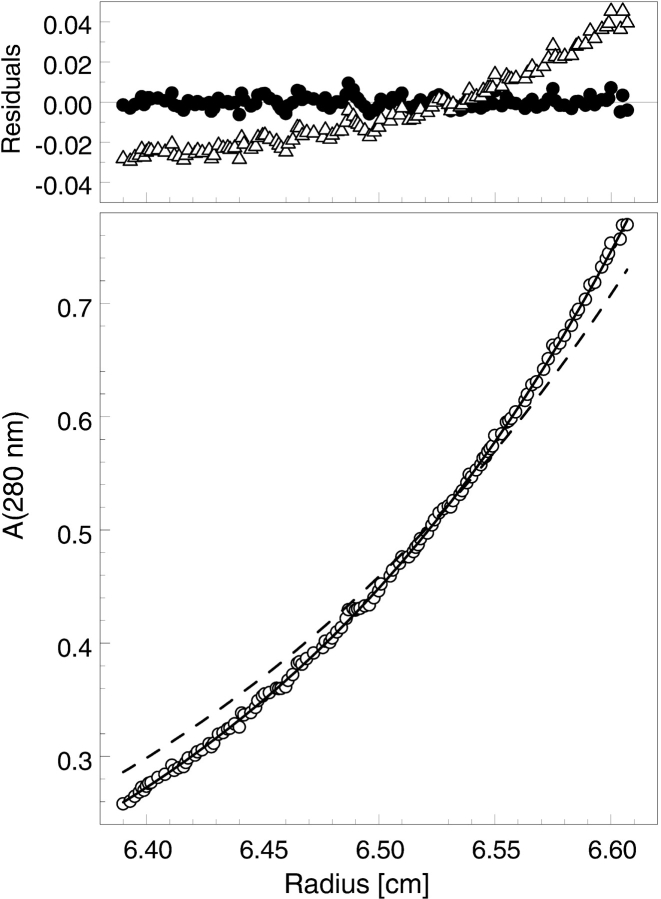
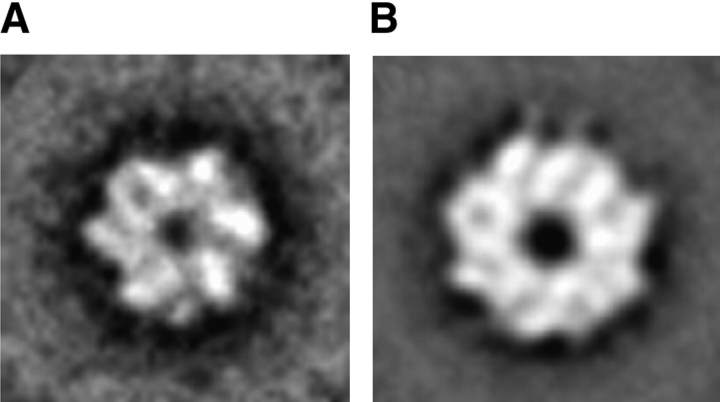
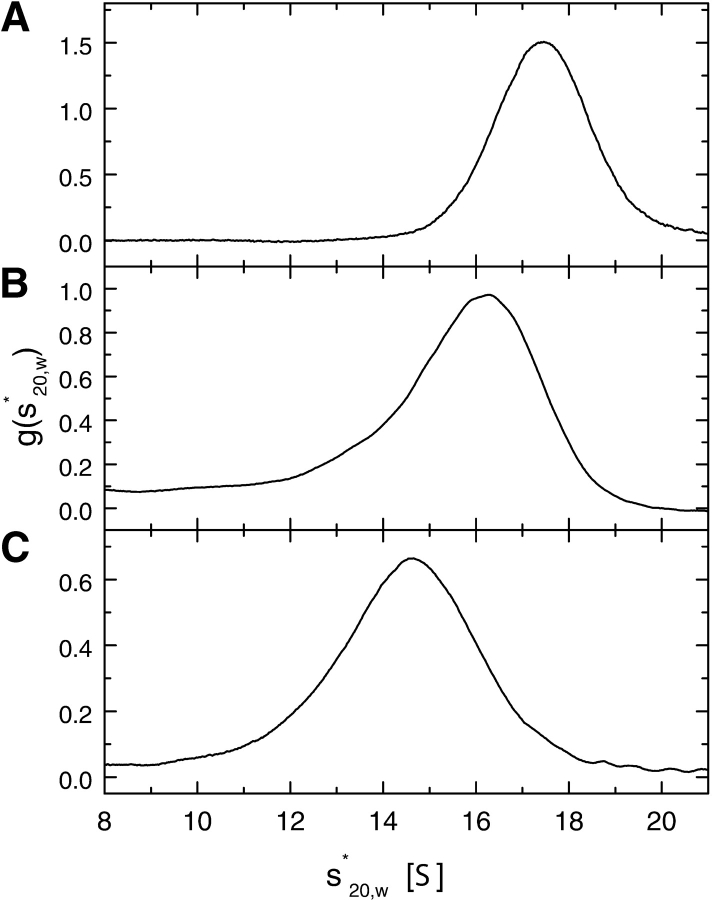
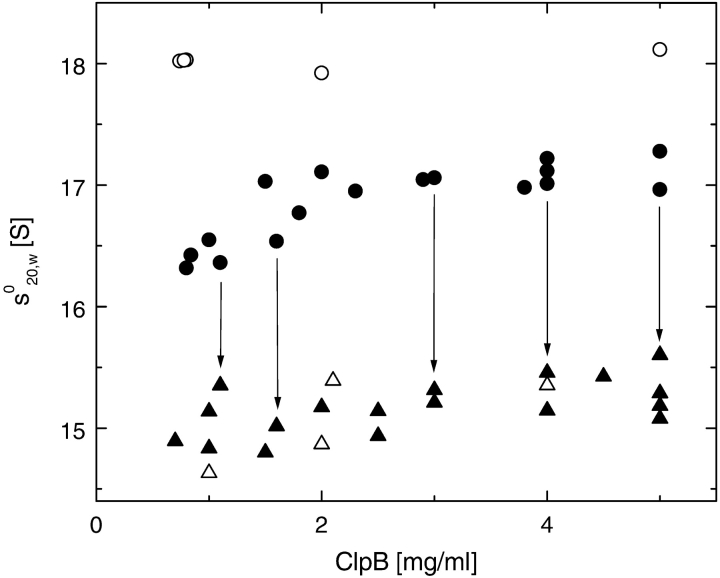
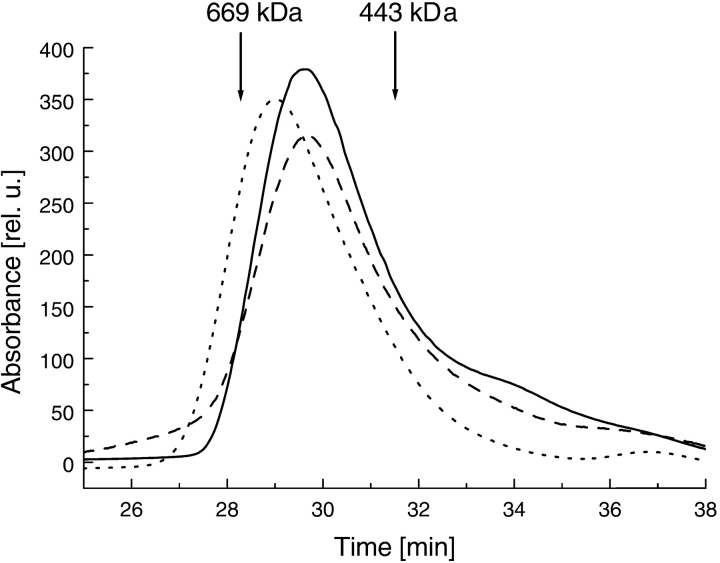
ClpB is a member of the bacterial protein-disaggregating chaperone machinery and belongs to the AAA(+) superfamily of ATPases associated with various cellular activities. The mechanism of ClpB-assisted reactivation of strongly aggregated proteins is unknown and the oligomeric state of ClpB has been under discussion. Sedimentation equilibrium and sedimentation velocity show that, under physiological ionic strength in the absence of nucleotides, ClpB from Escherichia coli undergoes reversible self-association that involves protein concentration-dependent populations of monomers, heptamers, and intermediate-size oligomers. Under low ionic strength conditions, a heptamer becomes the predominant form of ClpB. In contrast, ATP gamma S, a nonhydrolyzable ATP analog, as well as ADP stabilize hexameric ClpB. Consistently, electron microscopy reveals that ring-type oligomers of ClpB in the absence of nucleotides are larger than those in the presence of ATP gamma S. Thus, the binding of nucleotides without hydrolysis of ATP produces a significant change in the self-association equilibria of ClpB: from reactions supporting formation of a heptamer to those supporting a hexamer. Our results show how ClpB and possibly other related AAA(+) proteins can translate nucleotide binding into a major structural transformation and help explain why previously published electron micrographs of some AAA(+) ATPases detected both six- and sevenfold particle symmetry.
Figures
References
-
- Bochtler, M., Hartmann, C., Song, H.K., Bourenkov, G.P., Bartunik, H.D., and Huber, R. 2000. The structures of HsIU and the ATP-dependent protease HsIU-HsIV. Nature 403 800–805. - PubMed
-
- Glover, J.R. and Lindquist, S. 1998. Hsp104, Hsp70, and Hsp40: A novel chaperone system that rescues previously aggregated proteins. Cell 94 73–82. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
