

Does protein kinase R mediate TNF-alpha- and ceramide-induced increases in expression and activation of matrix metalloproteinases in articular cartilage by a novel mechanism?
- PMID: 14979937
- PMCID: PMC400415
- DOI: 10.1186/ar1024
Does protein kinase R mediate TNF-alpha- and ceramide-induced increases in expression and activation of matrix metalloproteinases in articular cartilage by a novel mechanism?
Abstract
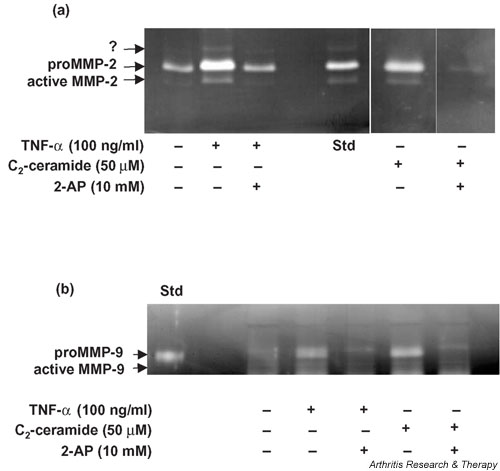
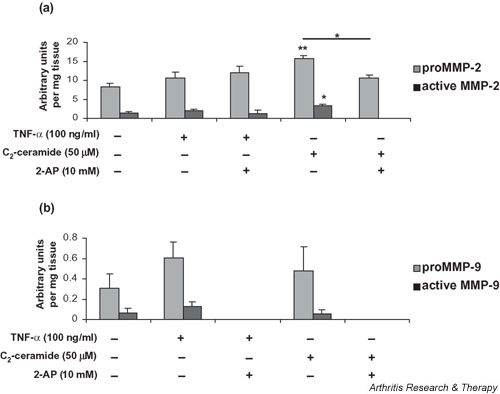
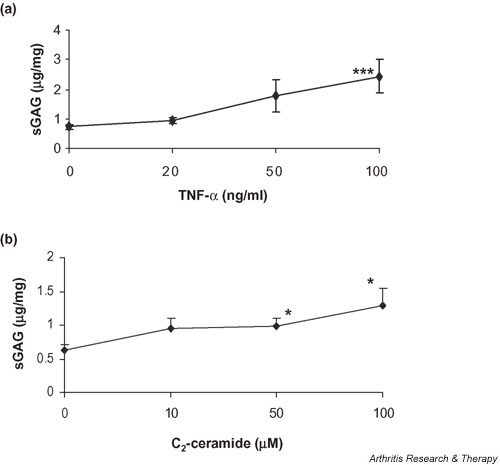
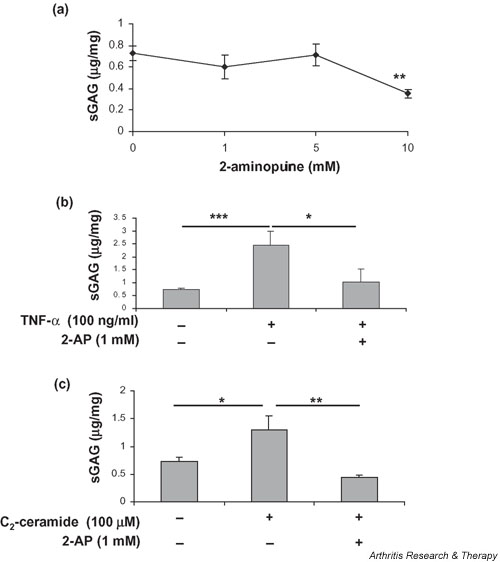
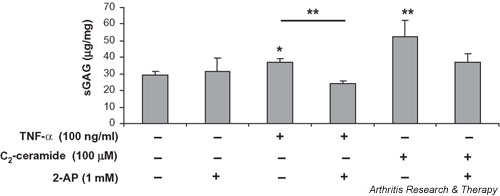
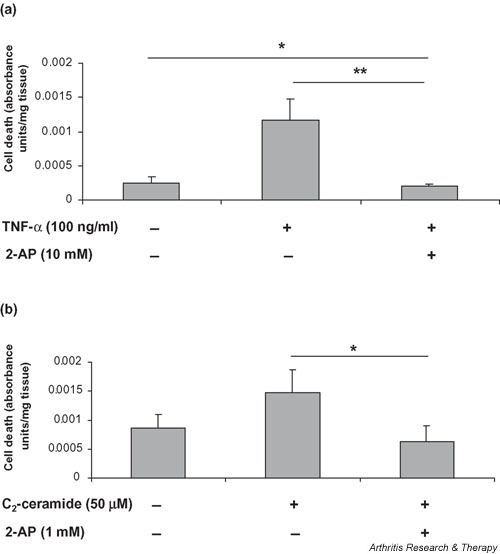
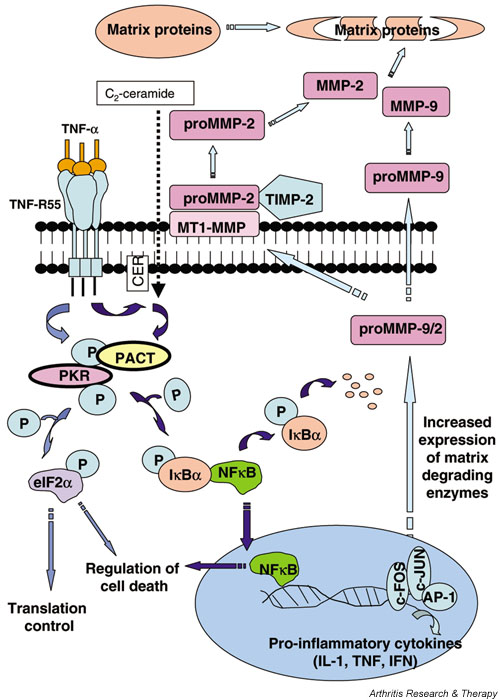
We investigated the role of the proinflammatory cytokine TNF-alpha, the second messenger C2-ceramide, and protein kinase R (PKR) in bovine articular cartilage degradation. Bovine articular cartilage explants were stimulated with C2-ceramide or TNF-alpha for 24 hours. To inhibit the activation of PKR, 2-aminopurine was added to duplicate cultures. Matrix metalloproteinase (MMP) expression and activation in the medium were analysed by gelatin zymography, proteoglycan release by the dimethylmethylene blue assay, and cell viability by the Cytotox 96(R) assay. C2-ceramide treatment of cartilage explants resulted in a significant release of both pro- and active MMP-2 into the medium. Small increases were also seen with TNF-alpha treatment. Incubation of explants with 2-aminopurine before TNF-alpha or C2-ceramide treatment resulted in a marked reduction in expression and activation of both MMP-2 and MMP-9. TNF-alpha and C2-ceramide significantly increased proteoglycan release into the medium, which was also inhibited by cotreatment with 2-aminopurine. A loss of cell viability was observed when explants were treated with TNF-alpha and C2-ceramide, which was found to be regulated by PKR. We have shown that C2-ceramide and TNF-alpha treatment of articular cartilage result in the increased synthesis and activation of MMPs, increased release of proteoglycan, and increased cell death. These effects are abrogated by treatment with the PKR inhibitor 2-aminopurine. Collectively, these results suggest a novel role for PKR in the synthesis and activation of MMPs and support our hypothesis that PKR and its activator, PACT, are implicated in the cartilage degradation that occurs in arthritic disease.
Figures
References
-
- Aigner T, Zien A, Gehrsitz A, Gebhard PM, McKenna L. Anabolic and catabolic gene expression pattern analysis in normal versus osteoarthritic cartilage using complementary DNA-array technology. Arthritis Rheum. 2001;44:2777–2789. doi: 10.1002/1529-0131(200112)44:12<2777::AID-ART465>3.0.CO;2-H. - DOI - PubMed
-
- Yoshihara Y, Nakamura H, Obata K, Yamada H, Hayakawa T, Fujikawa K, Okada Y. Matrix metalloproteinases and tissue inhibitors of metalloproteinases in synovial fluids from patients with rheumatoid arthritis or osteoarthritis. Ann Rheum Dis. 2000;59:455–461. doi: 10.1136/ard.59.6.455. - DOI - PMC - PubMed
-
- Goldring MB. Osteoarthritis and cartilage: The role of cytokines. Curr Rheumatol Rep. 2000;2:459–465. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
