

Structural basis of tyrosine sulfation and VH-gene usage in antibodies that recognize the HIV type 1 coreceptor-binding site on gp120
- PMID: 14981267
- PMCID: PMC365685
- DOI: 10.1073/pnas.0308527100
Structural basis of tyrosine sulfation and VH-gene usage in antibodies that recognize the HIV type 1 coreceptor-binding site on gp120
Abstract
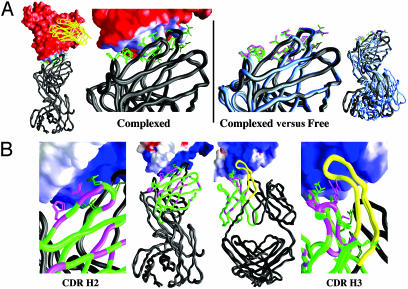
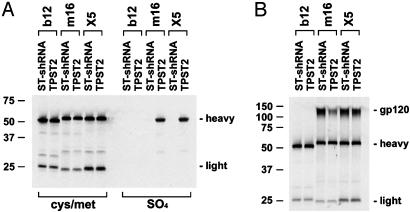
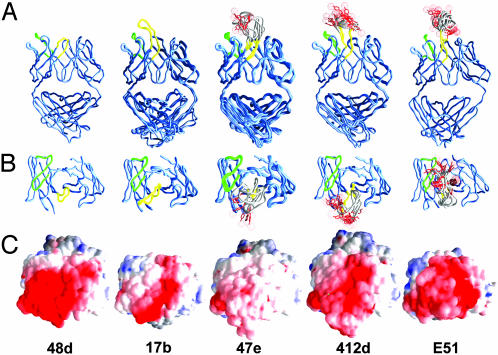
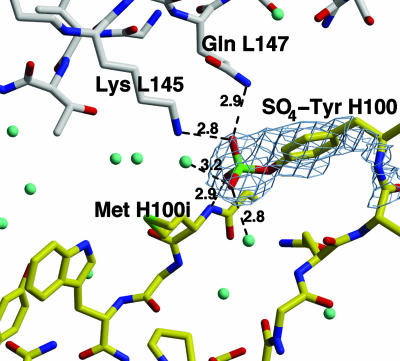
The conserved surface of the HIV-1 gp120 envelope glycoprotein that binds to the HIV-1 coreceptor is protected from humoral recognition by multiple layers of camouflage. Here we present sequence and genomic analyses for 12 antibodies that pierce these defenses and determine the crystal structures of 5. The data reveal mechanisms and atomic-level details for three unusual immune features: posttranslational mimicry of coreceptor by tyrosine sulfation of antibody, an alternative molecular mechanism controlling such sulfation, and highly selective V(H)-gene usage. When confronted by extraordinary viral defenses, the immune system unveils novel adaptive capabilities, with tyrosine sulfation enhancing the vocabulary of antigen recognition.
Figures

References
-
- Profy, A. T., Salinas, P. A., Eckler, L. I., Dunlop, N. M., Nara, P. L. & Putney, S. D. (1990) J. Immunol. 144, 4641-4647. - PubMed
-
- Wyatt, R., Kwong, P. D., Desjardins, E., Sweet, R. W., Robinson, J., Hendrickson, W. A. & Sodroski, J. (1998) Nature 393, 705-711. - PubMed
-
- Kwong, P. D., Doyle, M. L., Casper, D. J., Cicala, C., Leavitt, S. A., Majeed, S., Steenbeke, T. D., Venturi, M., Chaikin, I., Fung, M., et al. (2002) Nature 420, 678-682. - PubMed
-
- Wei, X., Decker, J. M., Wang, S., Hui, H., Kappes, J. C., Wu, X., Salazar-Gonzalez, J. F., Salazar, G. M., Kilby, M. J., Saag, M. S., et al. (2003) Nature 422, 307-312. - PubMed
-
- Barre-Sinoussi, F., Chermann, J. C., Rey, F., Nugeyre, M. T., Chamaret, S., Gruest, J., Dauguet, C., Axler-Blin, C., Vezinet-Brun, F., Rouzioux, C., et al. (1983) Science 220, 868-871. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
