

The solution structure of the N-terminal domain of E3L shows a tyrosine conformation that may explain its reduced affinity to Z-DNA in vitro
- PMID: 14981270
- PMCID: PMC365686
- DOI: 10.1073/pnas.0308612100
The solution structure of the N-terminal domain of E3L shows a tyrosine conformation that may explain its reduced affinity to Z-DNA in vitro
Abstract
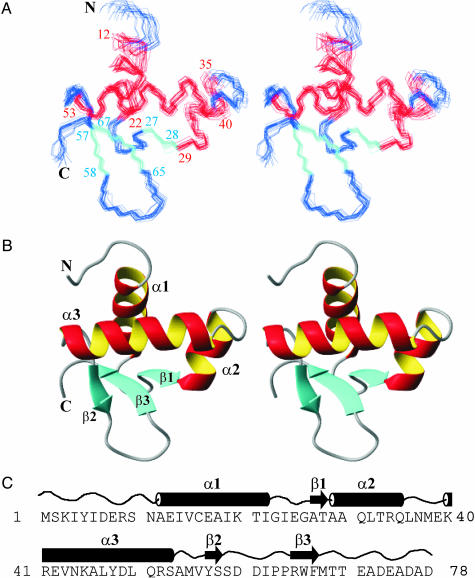
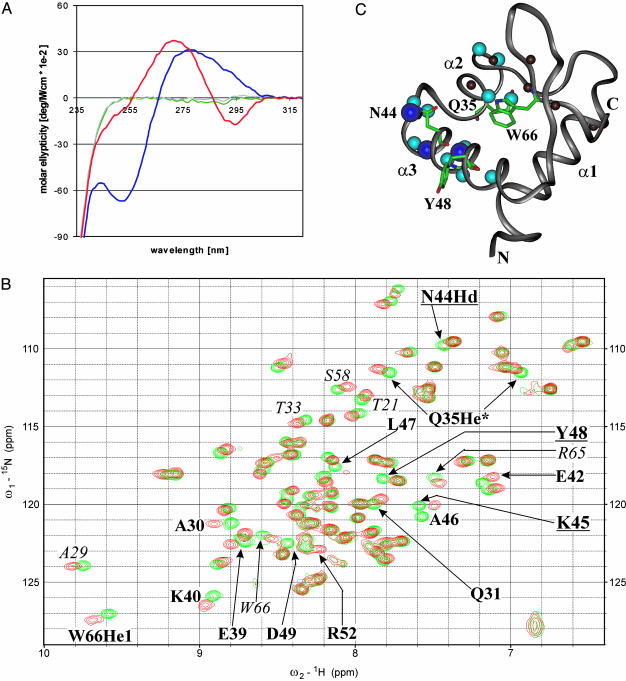
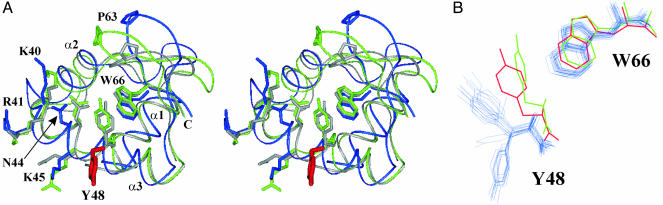
The N-terminal domain of the vaccinia virus protein E3L (Z alpha(E3L)) is essential for full viral pathogenicity in mice. It has sequence similarity to the high-affinity human Z-DNA-binding domains Z alpha(ADAR1) and Z alpha(DLM1). Here, we report the solution structure of Z alpha(E3L) and the chemical shift map of its interaction surface with Z-DNA. The global structure and the Z-DNA interaction surface of Z alpha(E3L) are very similar to the high-affinity Z-DNA-binding domains Z alpha(ADAR1) and Z alpha(DLM1). However, the key Z-DNA contacting residue Y48 of Z alpha(E3L) adopts a different side chain conformation in unbound Z alpha(E3L), which requires rearrangement for binding to Z-DNA. This difference suggests a molecular basis for the significantly lower in vitro affinity of Z alpha(E3L) to Z-DNA compared with its homologues.
Figures
References
-
- Cohen, J. (2001) Science 294, 985. - PubMed
-
- Kim, Y. G., Lowenhaupt, K., Oh, D. B., Kim, K. K. & Rich, A. (2004) Proc. Natl. Acad. Sci. USA, in press.
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
