

Miltefosine induces apoptosis-like death in Leishmania donovani promastigotes
- PMID: 14982775
- PMCID: PMC353131
- DOI: 10.1128/AAC.48.3.852-859.2004
Miltefosine induces apoptosis-like death in Leishmania donovani promastigotes
Abstract
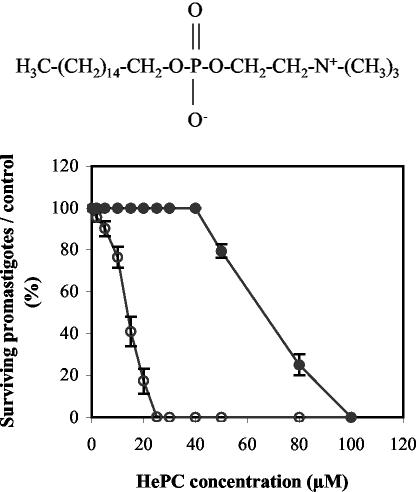
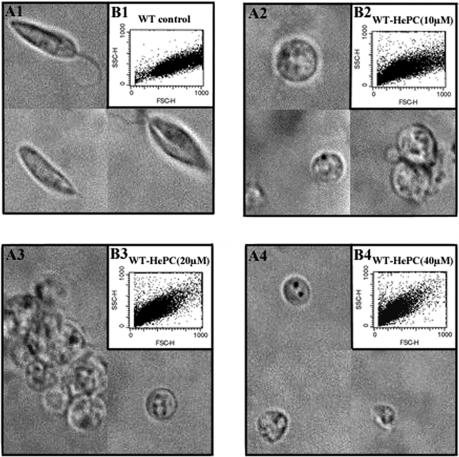
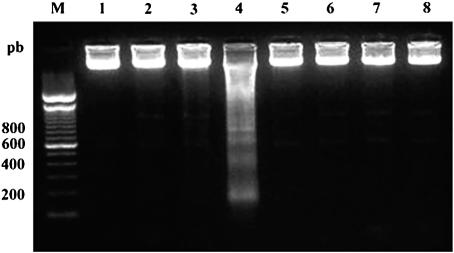
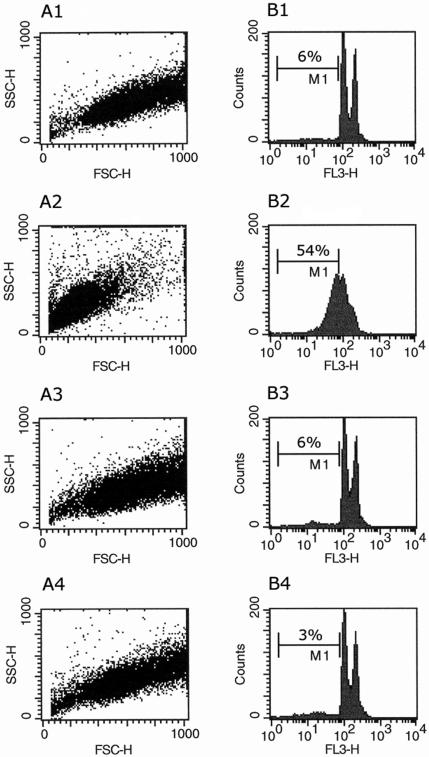
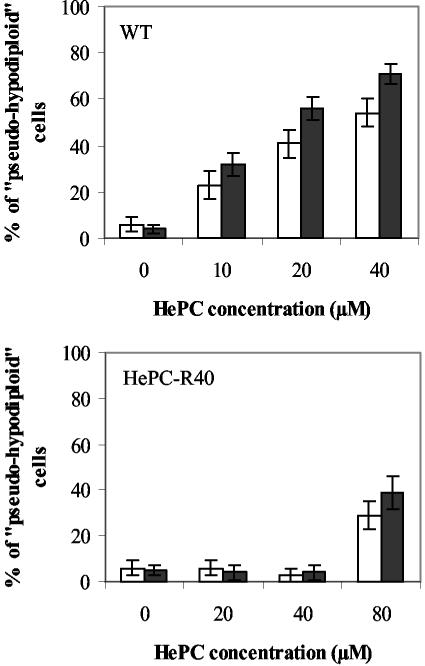
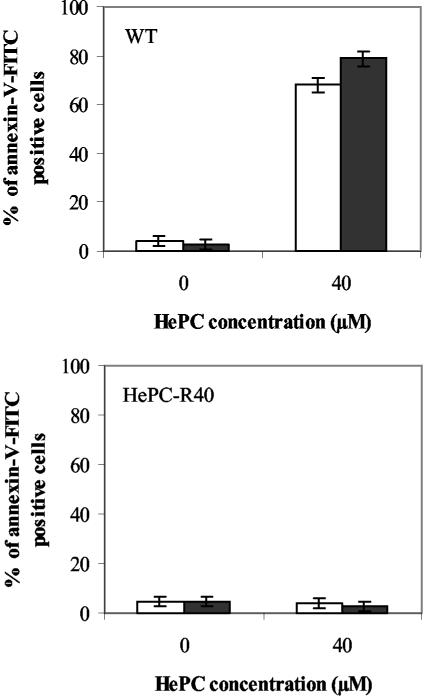
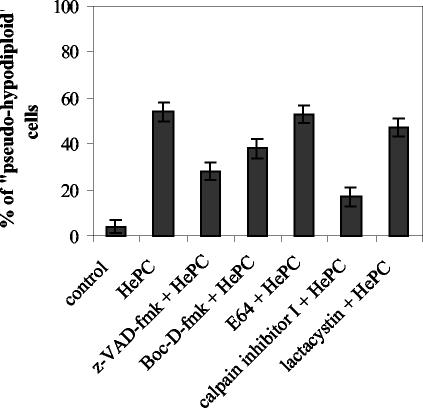
Miltefosine (hexadecylphosphocholine [HePC]) has proved to be a potent oral treatment for human visceral leishmaniasis due to Leishmania donovani. The molecular mechanisms that contribute to the antileishmanial activity of HePC are still unknown. We report that in wild-type promastigotes of Leishmania donovani HePC is able to induce a cell death process with numerous cytoplasmic, nuclear, and membrane features of metazoan apoptosis, including cell shrinkage, DNA fragmentation into oligonucleosome-sized fragments, and phosphatidylserine exposure. None of these changes were detected in an HePC-resistant clone treated with the same drug concentration. Therefore, HePC does not appear to kill L. donovani promastigotes by a direct toxic mechanism but, rather, kills the promastigotes by an indirect one. Pretreatment of wild-type promastigotes with two broad caspase inhibitors, z-Val-Ala-DL-Asp(methoxy)-fluoromethylketone and Boc-Asp(methoxy)-fluoromethylketone, as well as a broad protease inhibitor, calpain inhibitor I, prior to drug exposure interfered with DNA fragmentation but did not prevent cell shrinkage or phosphatidylserine externalization. These data suggest that at least part of the apoptotic machinery operating in wild-type promastigotes involves proteases. Identification of the death-signaling pathways activated in HePC-sensitive parasites appears to be essential for a better understanding of the molecular mechanisms of action and resistance in these parasites.
Figures
Similar articles
-
Membrane sterol depletion impairs miltefosine action in wild-type and miltefosine-resistant Leishmania donovani promastigotes.J Antimicrob Chemother. 2009 Nov;64(5):993-1001. doi: 10.1093/jac/dkp321. Epub 2009 Sep 12. J Antimicrob Chemother. 2009. PMID: 19749205
-
Miltefosine induces programmed cell death in Leishmania amazonensis promastigotes.Mem Inst Oswaldo Cruz. 2011 Jun;106(4):507-9. doi: 10.1590/s0074-02762011000400021. Mem Inst Oswaldo Cruz. 2011. PMID: 21739043
-
A mitochondrial HSP70 (HSPA9B) is linked to miltefosine resistance and stress response in Leishmania donovani.Parasit Vectors. 2016 Dec 1;9(1):621. doi: 10.1186/s13071-016-1904-8. Parasit Vectors. 2016. PMID: 27906059 Free PMC article.
-
Apoptosis and apoptotic mimicry: the Leishmania connection.Cell Mol Life Sci. 2010 May;67(10):1653-9. doi: 10.1007/s00018-010-0291-0. Epub 2010 Feb 7. Cell Mol Life Sci. 2010. PMID: 20140747 Free PMC article. Review.
-
Apoptosis and apoptotic mimicry in Leishmania: an evolutionary perspective.Front Cell Infect Microbiol. 2012 Jul 12;2:96. doi: 10.3389/fcimb.2012.00096. eCollection 2012. Front Cell Infect Microbiol. 2012. PMID: 22912937 Free PMC article. Review.
Cited by
-
Synthesis of new quinazolin-2,4-diones as anti-Leishmania mexicana agents.Mol Divers. 2016 Nov;20(4):821-828. doi: 10.1007/s11030-016-9693-8. Epub 2016 Aug 16. Mol Divers. 2016. PMID: 27531196
-
Evaluation of the Ability of Miltefosine Associated with Topical GM-CSF in Modulating the Immune Response of Patients with Cutaneous Leishmaniasis.J Immunol Res. 2020 Aug 6;2020:2789859. doi: 10.1155/2020/2789859. eCollection 2020. J Immunol Res. 2020. PMID: 32851099 Free PMC article.
-
Mitochondrial disruption and DNA fragmentation in Trypanosoma cruzi induced by naphthoimidazoles synthesized from beta-lapachone.Parasitol Res. 2007 Sep;101(4):895-905. doi: 10.1007/s00436-007-0556-1. Epub 2007 Jun 2. Parasitol Res. 2007. PMID: 17546464
-
Activities of naphthylisoquinoline alkaloids and synthetic analogs against Leishmania major.Antimicrob Agents Chemother. 2007 Jan;51(1):188-94. doi: 10.1128/AAC.00936-06. Epub 2006 Nov 6. Antimicrob Agents Chemother. 2007. PMID: 17088484 Free PMC article.
-
16alpha-Hydroxycleroda-3,13 (14)Z-dien-15,16-olide from Polyalthia longifolia: a safe and orally active antileishmanial agent.Br J Pharmacol. 2010 Mar;159(5):1143-50. doi: 10.1111/j.1476-5381.2009.00609.x. Epub 2010 Feb 5. Br J Pharmacol. 2010. PMID: 20136832 Free PMC article.
References
-
- Ameisen, J. C. 1995. Apoptosis in a unicellular eukaryotes (Trypanosoma cruzi): implications for the evolutionary origin and role of programmed cell in the control of cell proliferation, differentiation and survival. Cell Death Differ. 2:285-300. - PubMed
-
- Ameisen, J. C. 2002. On the origin, evolution, and nature of programmed cell death: a timeline of four billion years. Cell Death Differ. 9:367-393. - PubMed
-
- Aravind, L., V. M. Dixit, and E. V. Koonin. 2001. Apoptotic molecular machinery: vastly increased complexity in vertebrates revealed by genome comparisons. Science 291:1279-1284. - PubMed
-
- Arnoult, D., K. Akarid, A. Grodet, P. X. Petit, J. Estaquier, and J. C. Ameisen. 2002. On the evolution of programmed cell death: apoptosis of the unicellular eukaryote Leishmania major involves cysteine proteinase activation and mitochondrion permeabilization. Cell Death Differ. 9:65-81. - PubMed
-
- Boggs, K., C. O. Rock, and S. Jackowski. 1998. The antiproliferative effect of hexadecylphosphocholine toward HL60 cells is prevented by exogenous lysophosphatidylcholine. Biochim. Biophys. Acta 1389:1-12. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
