

Human pancreatic islet-derived progenitor cell engraftment in immunocompetent mice
- PMID: 14982836
- PMCID: PMC1613272
- DOI: 10.1016/S0002-9440(10)63170-7
Human pancreatic islet-derived progenitor cell engraftment in immunocompetent mice
Abstract
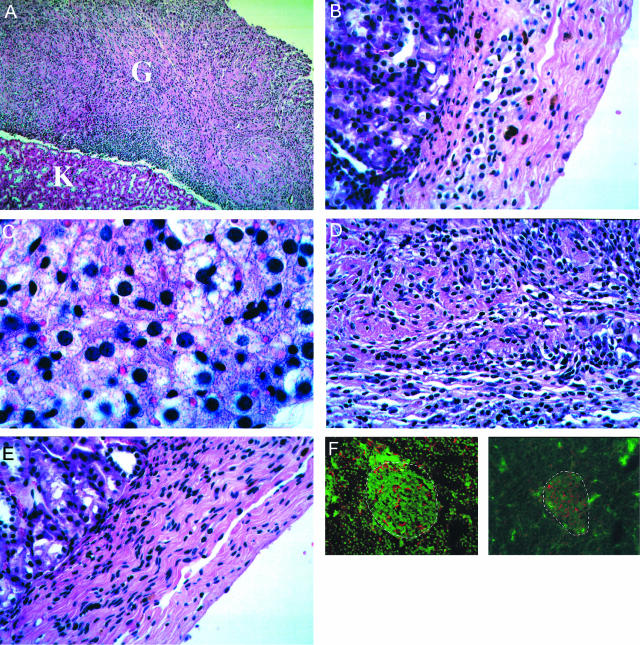
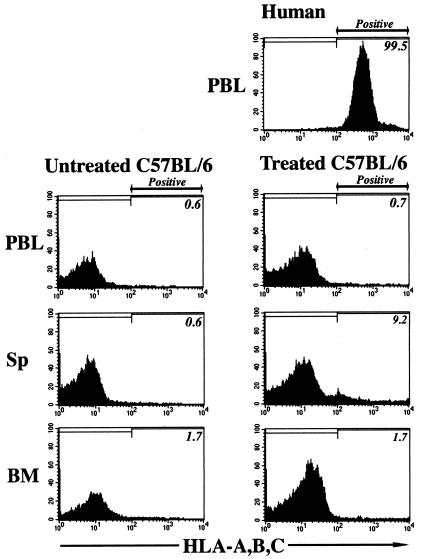
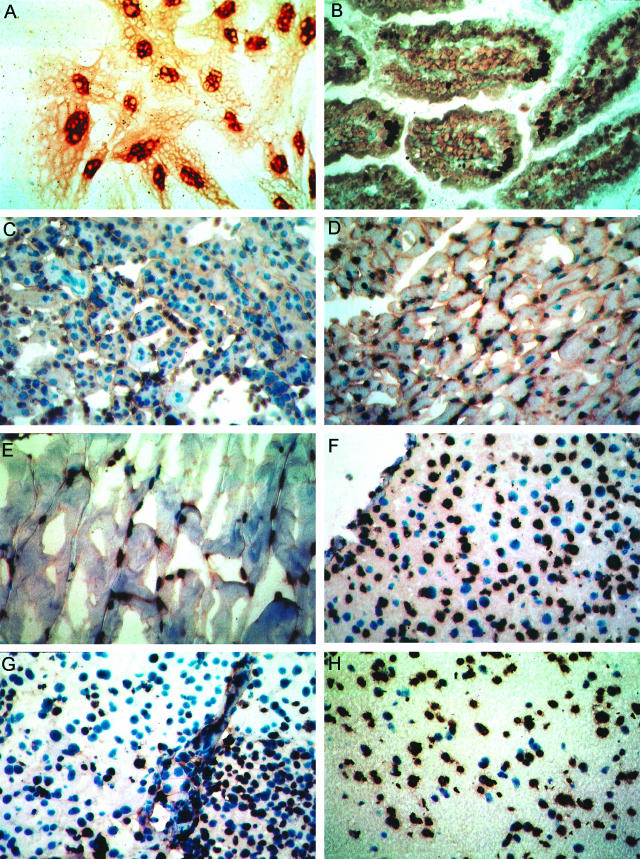
The potential for the use of stem/progenitor cells for the restoration of injured or diseased tissues has garnered much interest recently, establishing a new field of research called regenerative medicine. Attention has been focused on embryonic stem cells derived from human fetal tissues. However, the use of human fetal tissue for research and transplantation is controversial. An alternative is the isolation and utilization of multipotent stem/progenitor cells derived from adult donor tissues. We have previously reported on the isolation, propagation, and partial characterization of a population of stem/progenitor cells isolated from the pancreatic islets of Langerhans of adult human donor pancreata. Here we show that these human adult tissue-derived cells, nestin-positive islet-derived stem/progenitor cells, prepared from human adult pancreata survive engraftment and produce tissue chimerism when transplanted into immunocompetent mice either under the kidney capsule or by systemic injection. These xenografts seem to induce immune tolerance by establishing a mixed chimerism in the mice. We propose that a population of stem/progenitor cells isolated from the islets of the pancreas can cross xenogeneic transplantation immune barriers, induce tissue tolerance, and grow.
Figures







References
-
- Blau HM, Brazelton TR, Weimann JM. The evolving concept of a stem cell: entity or function? Cell. 2001;105:829–841. - PubMed
-
- Clarke DL, Johansson CB, Wilbertz J, Veress B, Nilsson E, Karlstrom H, Lendahl U, Frisen J. Generalized potential of adult neural stem cells. Science. 2000;288:1660–1663. - PubMed
-
- Gage FH. Mammalian neural stem cells. Science. 2000;287:1433–1438. - PubMed
-
- Jiang Y, Jahagirdar BN, Reinhardt RL, Schwartz RE, Keene CD, Ortiz-Gonzalez XR, Reyes M, Lenvik T, Lund T, Blackstad M, Du J, Aldrich S, Lisberg A, Low WC, Largaespada DA, Verfaillie CM. Pluripotency of mesenchymal stem cells derived from adult marrow. Nature. 2002;418:41–49. - PubMed
-
- Slack JM. Stem cells in epithelial tissues. Science. 2000;287:1431–1433. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
