

Heterodimeric interactions among the 1-amino-cyclopropane-1-carboxylate synthase polypeptides encoded by the Arabidopsis gene family
- PMID: 14983000
- PMCID: PMC356941
- DOI: 10.1073/pnas.0308515101
Heterodimeric interactions among the 1-amino-cyclopropane-1-carboxylate synthase polypeptides encoded by the Arabidopsis gene family
Abstract
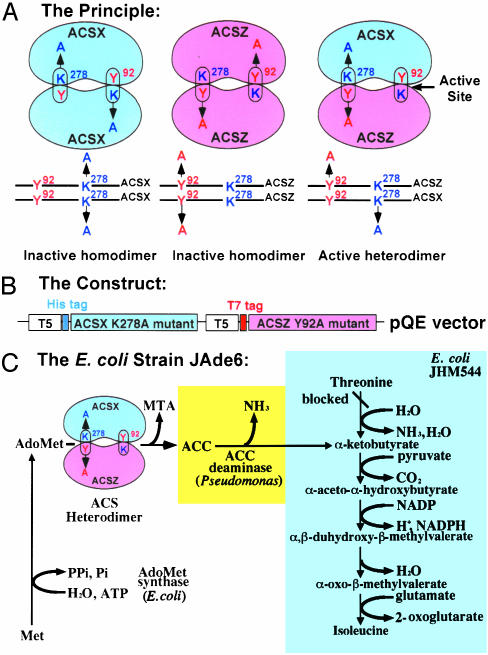
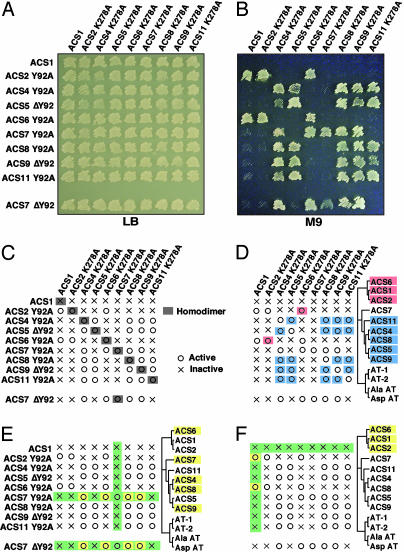
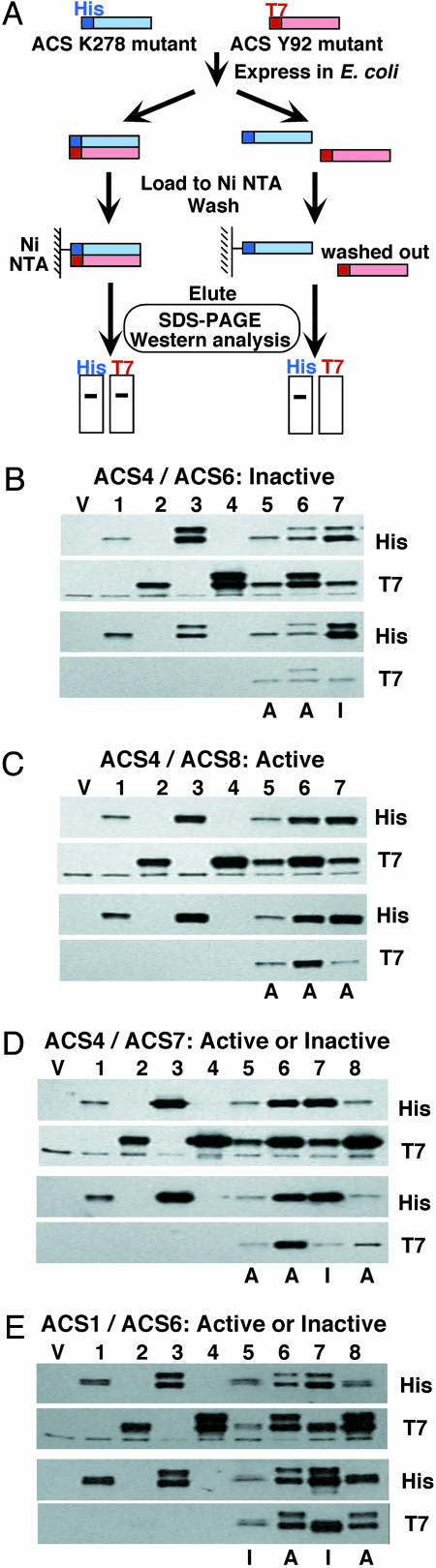
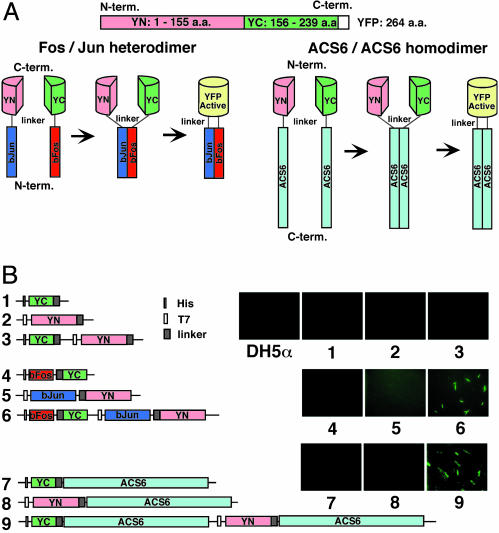
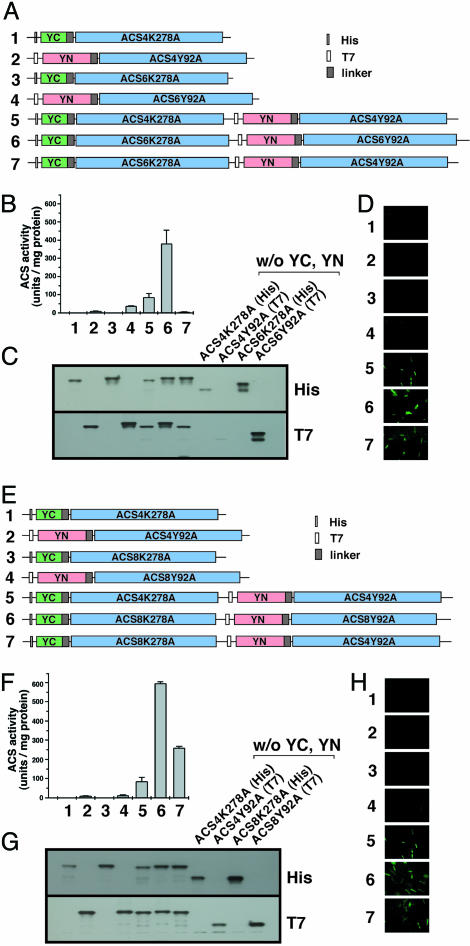
The pyridoxal phosphate-dependent enzyme, 1-aminocyclopropane-1-carboxylate synthase (ACS; EC 4.4.1.14), catalyzes the rate-limiting step in the ethylene biosynthetic pathway in plants. The Arabidopsis genome encodes nine ACS polypeptides that form eight functional (ACS2, ACS4-9, ACS11) and one nonfunctional (ACS1) homodimers. Because the enzyme is a homodimer with shared active sites, the question arises whether the various polypeptides can form functional heterodimers. Intermolecular complementation experiments in Escherichia coli by coexpressing the K278A and Y92A mutants of different polypeptides show that all of them have the capacity to heterodimerize. However, functional heterodimers are formed only among gene family members that belong to one or the other of the two phylogenetic branches. ACS7 is an exception to this rule, which forms functional heterodimers with some members of both branches when it provides the wt K278 residue. ACS1, the nonfunctional polypeptide as a homodimer, can also form functional heterodimers with members of its phylogenetic branch when its partners provide the wt K278 residue. The ACS gene family products can potentially form 45 homo- and heterodimers of which 25 are functional. Bimolecular fluorescence complementation and biochemical coaffinity purification assays show that the inactivity of certain heterodimers is not due to the absence of heterodimerization but rather to structural restraint(s) that prevents the shared active sites from being functional. We propose that functional heterodimerization enhances the isozyme diversity of the ACS gene family and provides physiological versatility by being able to operate in a broad gradient of S-adenosylmethionine concentration in various cells/tissues during plant growth and development. Nonfunctional heterodimerization may also play a regulatory role during the plant life cycle.
Figures
Similar articles
-
Unique and overlapping expression patterns among the Arabidopsis 1-amino-cyclopropane-1-carboxylate synthase gene family members.Plant Physiol. 2004 Oct;136(2):2982-3000. doi: 10.1104/pp.104.049999. Epub 2004 Oct 1. Plant Physiol. 2004. PMID: 15466221 Free PMC article.
-
A combinatorial interplay among the 1-aminocyclopropane-1-carboxylate isoforms regulates ethylene biosynthesis in Arabidopsis thaliana.Genetics. 2009 Nov;183(3):979-1003. doi: 10.1534/genetics.109.107102. Epub 2009 Sep 14. Genetics. 2009. PMID: 19752216 Free PMC article.
-
Complementation analysis of mutants of 1-aminocyclopropane- 1-carboxylate synthase reveals the enzyme is a dimer with shared active sites.J Biol Chem. 1998 May 15;273(20):12509-14. doi: 10.1074/jbc.273.20.12509. J Biol Chem. 1998. PMID: 9575209
-
Biochemical diversity among the 1-amino-cyclopropane-1-carboxylate synthase isozymes encoded by the Arabidopsis gene family.J Biol Chem. 2003 Dec 5;278(49):49102-12. doi: 10.1074/jbc.M308297200. Epub 2003 Sep 10. J Biol Chem. 2003. PMID: 12968022
-
The Arabidopsis RING-type E3 ligase XBAT32 mediates the proteasomal degradation of the ethylene biosynthetic enzyme, 1-aminocyclopropane-1-carboxylate synthase 7.Plant J. 2012 Jul;71(1):23-34. doi: 10.1111/j.1365-313X.2012.04965.x. Epub 2012 Apr 17. Plant J. 2012. PMID: 22339729
Cited by
-
Ethylene in mutualistic symbioses.Plant Signal Behav. 2012 Dec;7(12):1634-8. doi: 10.4161/psb.22471. Epub 2012 Oct 16. Plant Signal Behav. 2012. PMID: 23072986 Free PMC article. Review.
-
Functional genomic analysis of the AUXIN RESPONSE FACTOR gene family members in Arabidopsis thaliana: unique and overlapping functions of ARF7 and ARF19.Plant Cell. 2005 Feb;17(2):444-63. doi: 10.1105/tpc.104.028316. Epub 2005 Jan 19. Plant Cell. 2005. PMID: 15659631 Free PMC article.
-
The RING-type E3 ligase RIE1 sustains leaf longevity by specifically targeting AtACS7 to fine-tune ethylene production in Arabidopsis.Proc Natl Acad Sci U S A. 2024 Nov 26;121(48):e2411271121. doi: 10.1073/pnas.2411271121. Epub 2024 Nov 20. Proc Natl Acad Sci U S A. 2024. PMID: 39565318 Free PMC article.
-
Visualization of molecular interactions by fluorescence complementation.Nat Rev Mol Cell Biol. 2006 Jun;7(6):449-56. doi: 10.1038/nrm1929. Nat Rev Mol Cell Biol. 2006. PMID: 16625152 Free PMC article. Review.
-
Regulation of ACS protein stability by cytokinin and brassinosteroid.Plant J. 2009 Feb;57(4):606-14. doi: 10.1111/j.1365-313X.2008.03711.x. Epub 2008 Oct 10. Plant J. 2009. PMID: 18980656 Free PMC article.
References
-
- Abeles, F. B., Morgan, P. W. & Saltveit, M. E., Jr. (1992) Ethylene in Plant Biology (Academic, San Diego).
-
- Yang, S. F. & Hoffman, N. E. (1984) Annu. Rev. Plant Physiol. 35, 155–189.
-
- Bleecker, A. B. & Kende, H. (2000) Annu. Rev. Cell Dev. Biol. 16, 1–18. - PubMed
-
- Liang, X.-W., Shen, N. F. & Theologis, A. (1996) Plant J. 10, 1027–1036. - PubMed
-
- Thomma, B. P. H. J., Penninckx, I. A. M. A., Broekaert, W. F. & Cammue, B. P. A. (2001) Curr. Opin. Immunol. 13, 63–68. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
