

Adiponectin-induced antiangiogenesis and antitumor activity involve caspase-mediated endothelial cell apoptosis
- PMID: 14983034
- PMCID: PMC356975
- DOI: 10.1073/pnas.0308671100
Adiponectin-induced antiangiogenesis and antitumor activity involve caspase-mediated endothelial cell apoptosis
Abstract
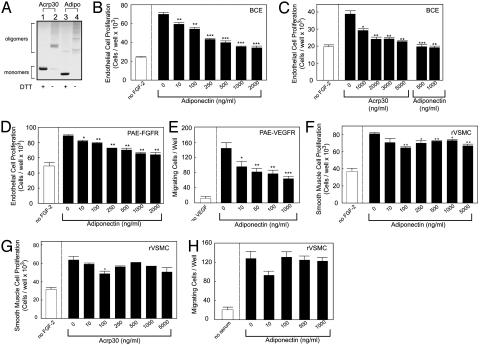
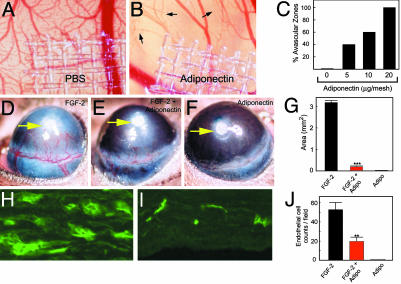
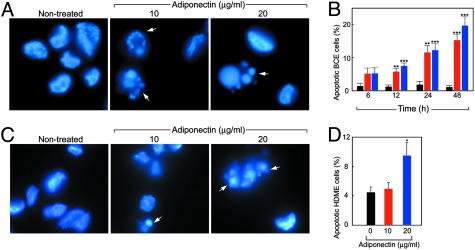
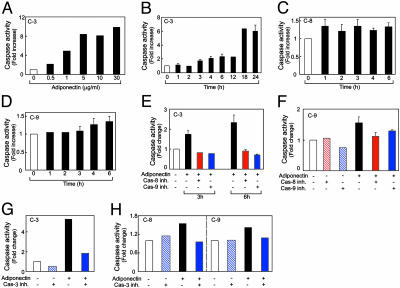
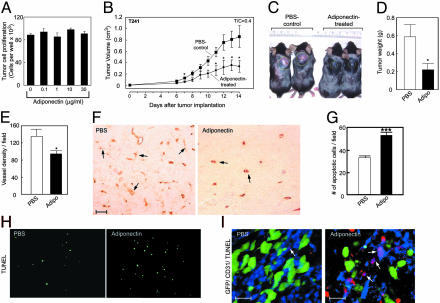
Obesity is a risk factor for the development of many severe human diseases such as cardiovascular disorders, diabetes, and cancer, which are tightly linked to angiogenesis. The adipose tissue produces several growth factors/hormones including leptin, tumor necrosis factor alpha, and adiponectin. It has been found that adiponectin levels are reduced in obesity. Here, we report a unique function of adiponectin as a negative regulator of angiogenesis. In vitro, adiponectin potently inhibits endothelial cell proliferation and migration. In the chick chorioallantoic membrane and the mouse corneal angiogenesis assays, adiponectin remarkably prevents new blood vessel growth. Further, we demonstrate that the antiendothelial mechanisms involve activation of caspase-mediated endothelial cell apoptosis. Adiponectin induces a cascade activation of caspases-8, -9, and -3, which leads to cell death. In a mouse tumor model, adiponectin significantly inhibits primary tumor growth. Impaired tumor growth is associated with decreased neovascularization, leading to significantly increased tumor cell apoptosis. These data demonstrate induction of endothelial apoptosis as an unique mechanism of adiponectin-induced antiangiogenesis. Adiponectin, as a direct endogenous angiogenesis inhibitor, may have therapeutic implications in the treatment of angiogenesis-dependent diseases.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
