

Disease progression despite early loss of polyglutamine protein expression in SCA7 mouse model
- PMID: 14985428
- PMCID: PMC6730412
- DOI: 10.1523/JNEUROSCI.4407-03.2004
Disease progression despite early loss of polyglutamine protein expression in SCA7 mouse model
Abstract
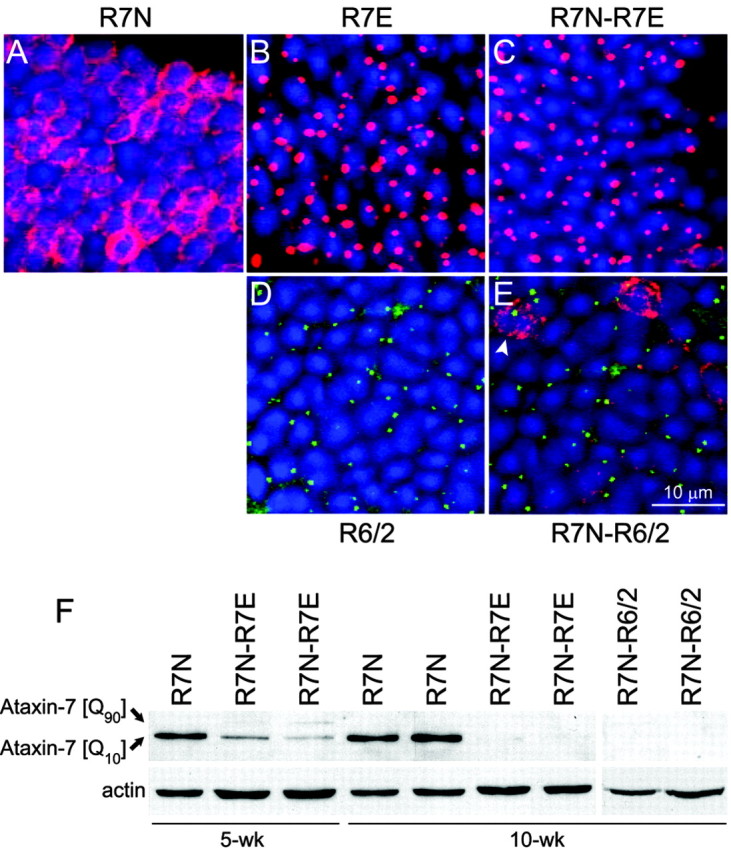
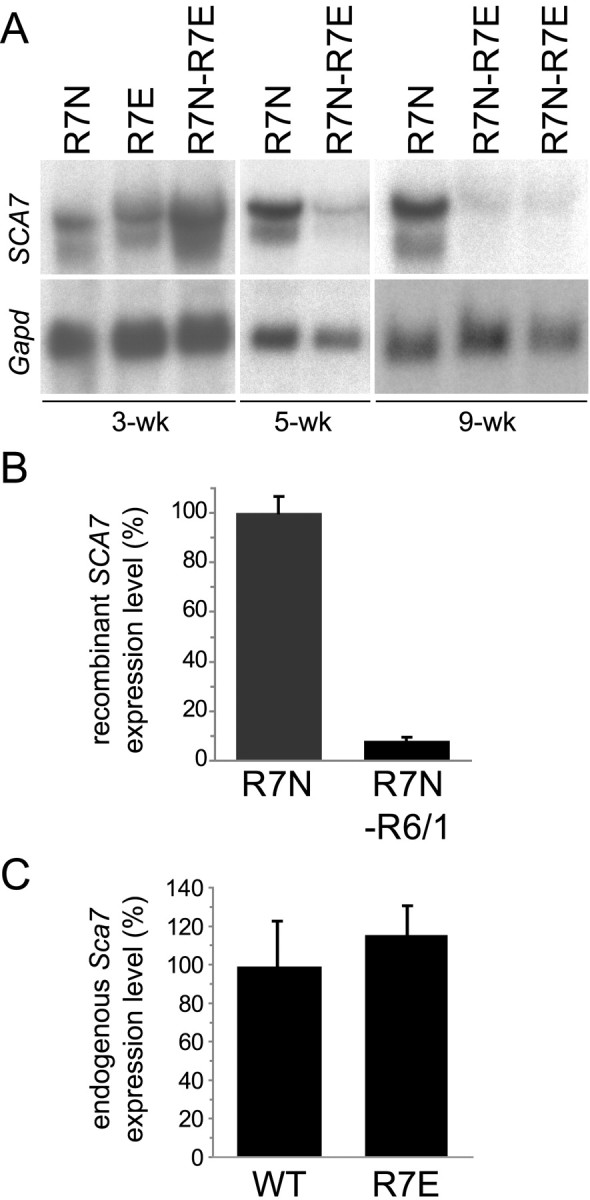
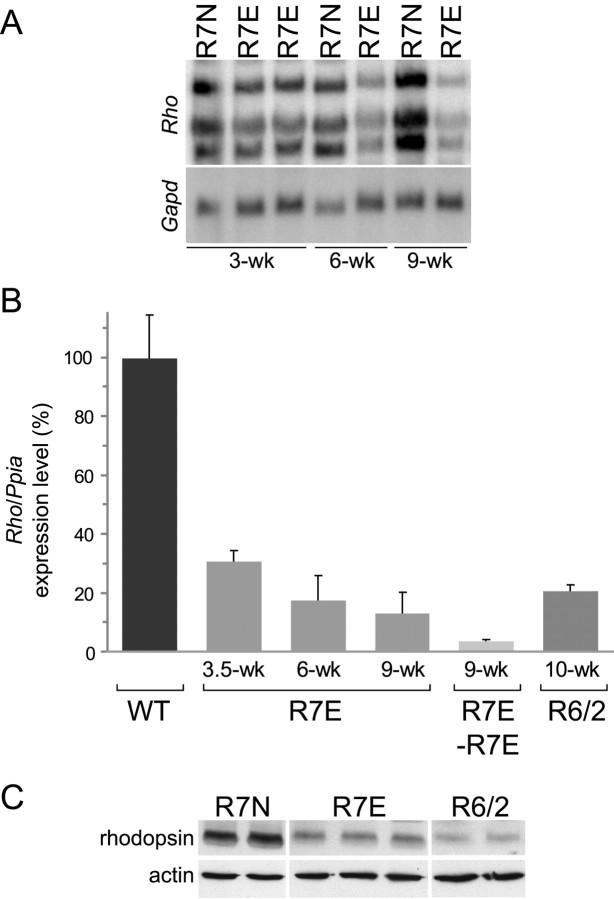
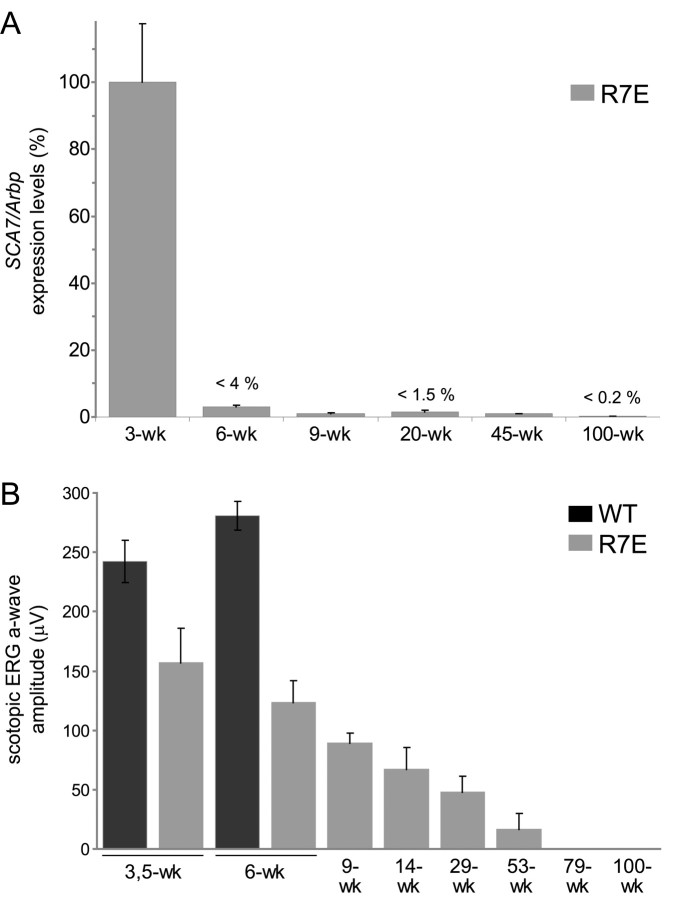
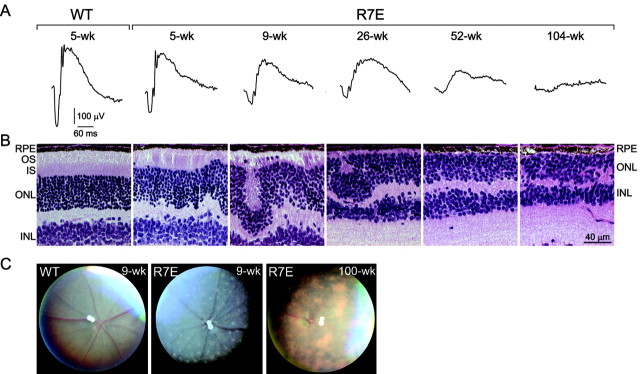
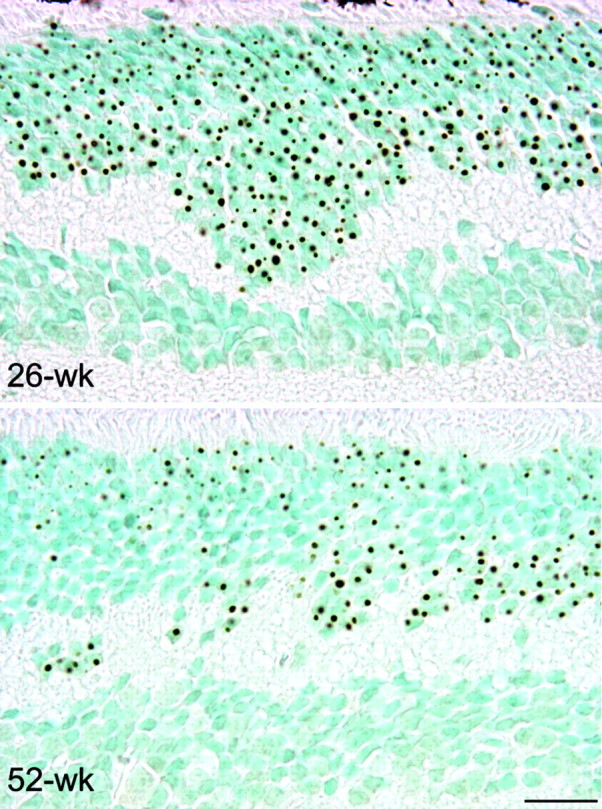
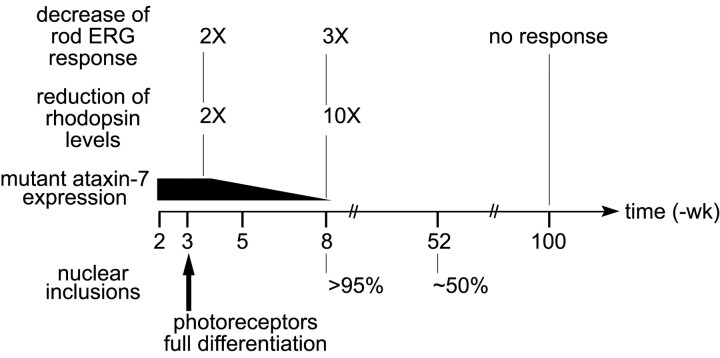
Nine neurodegenerative diseases including Huntington's disease (HD) and spinocerebellar ataxia type 7 (SCA7) are caused by an expansion of a polyglutamine (polyQ) stretch in the respective proteins. Aggregation of expanded polyQ-containing proteins into the nucleus is a hallmark of these diseases. Recent evidence indicates that transcriptional dysregulation may contribute to the molecular pathogenesis of these diseases. Using SCA7 and HD mouse models in which we recently described a retinal phenotype, we investigated whether altered gene expression underlies photoreceptor dysfunction. In both models, rhodopsin promoter activity was early and dramatically repressed, suggesting that downregulation of photoreceptor-specific genes plays a major role in polyQ-induced retinal dysfunction. Because the rhodopsin promoter drives mutant ataxin-7 expression in our SCA7 mice, we also assessed whether downregulation of mutant SCA7 transgene would reverse retinopathy progression and aggregate formation. Although residual expression of mutant ataxin-7 was found negligible from 9 weeks of age, SCA7 transgenic mice showed a progressive decline of photoreceptor activity leading to a complete loss of electroretinographic responses from 1 year of age. At this age, aggregates were cleared in only half of the photoreceptors, indicating that their formation is not fully reversible in this model. We demonstrate here that abolishing full-length mutant ataxin-7 expression did not reverse retinopathy progression in SCA7 mice, raising the possibility that some polyQ-induced pathological events might be irreversible.
Figures
Similar articles
-
Polyglutamine-expanded ataxin-7 antagonizes CRX function and induces cone-rod dystrophy in a mouse model of SCA7.Neuron. 2001 Sep 27;31(6):913-27. doi: 10.1016/s0896-6273(01)00422-6. Neuron. 2001. PMID: 11580893
-
Reduction of mutant ataxin-7 expression restores motor function and prevents cerebellar synaptic reorganization in a conditional mouse model of SCA7.Hum Mol Genet. 2013 Mar 1;22(5):890-903. doi: 10.1093/hmg/dds495. Epub 2012 Nov 29. Hum Mol Genet. 2013. PMID: 23197655 Free PMC article.
-
Antisense oligonucleotides targeting mutant Ataxin-7 restore visual function in a mouse model of spinocerebellar ataxia type 7.Sci Transl Med. 2018 Oct 31;10(465):eaap8677. doi: 10.1126/scitranslmed.aap8677. Sci Transl Med. 2018. PMID: 30381411 Free PMC article.
-
Molecular pathogenesis and cellular pathology of spinocerebellar ataxia type 7 neurodegeneration.Cerebellum. 2008;7(2):138-49. doi: 10.1007/s12311-008-0027-y. Cerebellum. 2008. PMID: 18418675 Free PMC article. Review.
-
Insights into the molecular basis of polyglutamine neurodegeneration from studies of a spinocerebellar ataxia type 7 mouse model.Cytogenet Genome Res. 2003;100(1-4):164-74. doi: 10.1159/000072851. Cytogenet Genome Res. 2003. PMID: 14526177 Review.
Cited by
-
PML clastosomes prevent nuclear accumulation of mutant ataxin-7 and other polyglutamine proteins.J Cell Biol. 2006 Jul 3;174(1):65-76. doi: 10.1083/jcb.200511045. J Cell Biol. 2006. PMID: 16818720 Free PMC article.
-
Altered Metabolic Signaling and Potential Therapies in Polyglutamine Diseases.Metabolites. 2024 May 31;14(6):320. doi: 10.3390/metabo14060320. Metabolites. 2024. PMID: 38921455 Free PMC article. Review.
-
Preventing polyglutamine-induced activation of c-Jun delays neuronal dysfunction in a mouse model of SCA7 retinopathy.Neurobiol Dis. 2007 Mar;25(3):571-81. doi: 10.1016/j.nbd.2006.11.002. Epub 2006 Dec 26. Neurobiol Dis. 2007. PMID: 17189700 Free PMC article.
-
Nonallele specific silencing of ataxin-7 improves disease phenotypes in a mouse model of SCA7.Mol Ther. 2014 Sep;22(9):1635-42. doi: 10.1038/mt.2014.108. Epub 2014 Jun 16. Mol Ther. 2014. PMID: 24930601 Free PMC article.
-
Glutamine-expanded ataxin-7 alters TFTC/STAGA recruitment and chromatin structure leading to photoreceptor dysfunction.PLoS Biol. 2006 Mar;4(3):e67. doi: 10.1371/journal.pbio.0040067. Epub 2006 Feb 28. PLoS Biol. 2006. PMID: 16494529 Free PMC article.
References
-
- Caplen NJ, Taylor JP, Statham VS, Tanaka F, Fire A, Morgan RA (2002) Rescue of polyglutamine-mediated cytotoxicity by double-stranded RNA-mediated RNA interference. Hum Mol Genet 11: 175-184. - PubMed
-
- Cattaneo E, Rigamonti D, Goffredo D, Zuccato C, Squitieri F, Sipione S (2001) Loss of normal huntingtin function: new developments in Huntington's disease research. Trends Neurosci 24: 182-188. - PubMed
-
- Chen S, Wang QL, Nie Z, Sun H, Lennon G, Copeland NG, Gilbert DJ, Jenkins NA, Zack DJ (1997) Crx, a novel Otx-like paired-homeodomain protein, binds to and transactivates photoreceptor cell-specific genes. Neuron 19: 1017-1030. - PubMed
-
- Davies SW, Turmaine M, Cozens BA, Difiglia M, Sharp AH, Ross CA, Scherzinger E, Wanker EE, Mangiarini L, Bates GP (1997) Formation of neuronal intranuclear inclusions underlies the neurological dysfunction in mice transgenic for the HD mutation. Cell 90: 537-548. - PubMed
-
- Furukawa T, Morrow EM, Cepko CL (1997) Crx, a novel otx-like homeobox gene, shows photoreceptor-specific expression and regulates photoreceptor differentiation. Cell 91: 531-541. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases