

Inhibition of different Lassa virus strains by alpha and gamma interferons and comparison with a less pathogenic arenavirus
- PMID: 14990737
- PMCID: PMC353741
- DOI: 10.1128/jvi.78.6.3162-3169.2004
Inhibition of different Lassa virus strains by alpha and gamma interferons and comparison with a less pathogenic arenavirus
Abstract
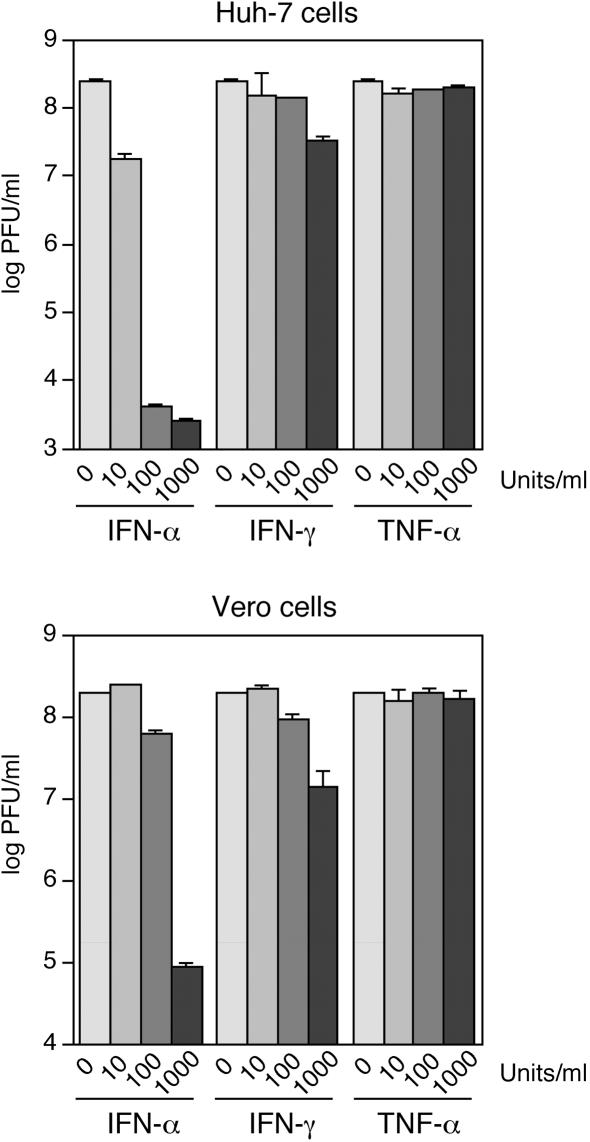
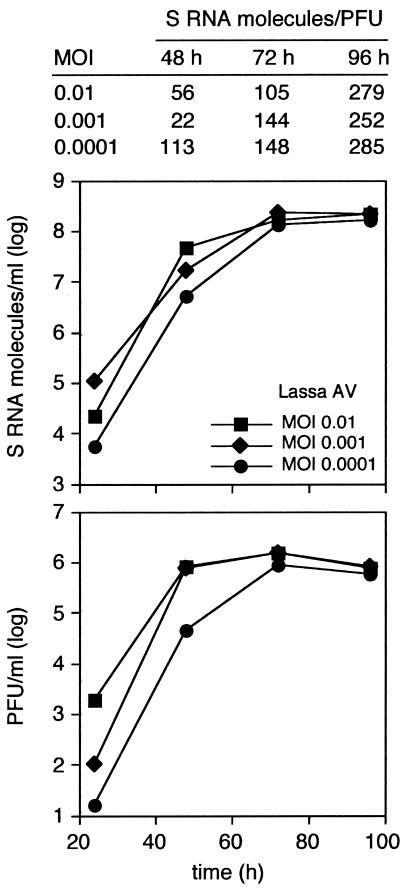
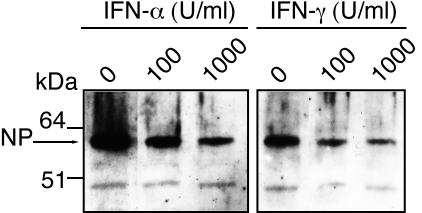
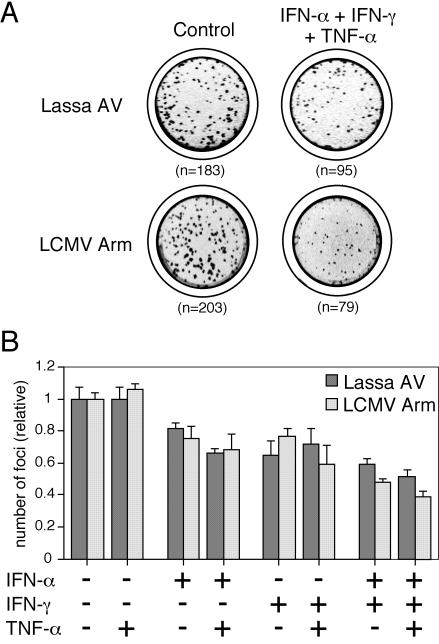
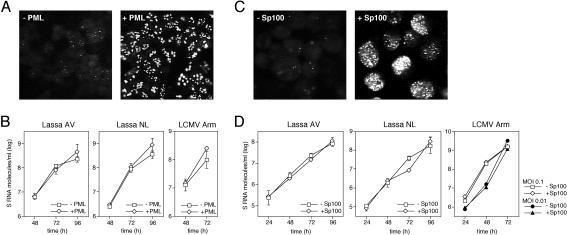
The high pathogenicity of Lassa virus is assumed to involve resistance to the effects of interferon (IFN). We have analyzed the effects of alpha IFN (IFN-alpha), IFN-gamma, and tumor necrosis factor alpha (TNF-alpha) on replication of Lassa virus compared to the related, but less pathogenic, lymphocytic choriomeningitis virus (LCMV). Three low-passage Lassa virus strains (AV, NL, and CSF), isolated from humans with mild to fulminant Lassa fever, were tested. Lassa virus replication was inhibited by IFN-alpha and IFN-gamma, but not TNF-alpha, in Huh7 and Vero cells. The degree of IFN sensitivity of a Lassa virus isolate did not correlate with disease severity in human patients. Furthermore, cytokine effects observed for Lassa virus and LCMV (strains CH-5692, Armstrong, and WE) were similar. To address the mechanisms involved in the IFN effect, we used cell lines in which overexpression of IFN-stimulated proteins promyelocytic leukemia protein (PML) and Sp100 could be induced. Both proteins reside in PML bodies, a cellular target of the LCMV and Lassa virus Z proteins. Overexpression of PML or Sp100 did not affect replication of either virus. This, together with the previous finding that PML knockout facilitates LCMV replication in vitro and in vivo (M. Djavani, J. Rodas, I. S. Lukashevich, D. Horejsh, P. P. Pandolfi, K. L. Borden, and M. S. Salvato, J. Virol. 75:6204-6208, 2001; W. V. Bonilla, D. D. Pinschewer, P. Klenerman, V. Rousson, M. Gaboli, P. P. Pandolfi, R. M. Zinkernagel, M. S. Salvato, and H. Hengartner, J. Virol. 76:3810-3818, 2002), describes PML as a mediator within the antiviral pathway rather than as a direct effector protein. In conclusion, the high pathogenicity of Lassa virus compared to LCMV is probably not due to increased resistance to the effects of IFN-alpha or IFN-gamma. Both cytokines inhibit replication which is relevant for the design of antiviral strategies against Lassa fever with the aim of enhancing the IFN response.
Figures
References
-
- Armstrong, C., and R. D. Lillie. 1934. Experimental lymphocytic choriomeningitis of monkeys and mice produced by a virus encountered in studies of the 1933 St. Louis encephalitis epidemic. Public Health Rep. 49:1019-1027.
-
- Asper, M., P. Hofmann, C. Osmann, J. Funk, C. Metzger, M. Bruns, F. J. Kaup, H. Schmitz, and S. Günther. 2001. First outbreak of callitrichid hepatitis in Germany: genetic characterization of the causative lymphocytic choriomeningitis virus strains. Virology 284:203-213. - PubMed
-
- Battegay, M., S. Cooper, A. Althage, J. Banziger, H. Hengartner, and R. M. Zinkernagel. 1991. Quantification of lymphocytic choriomeningitis virus with an immunological focus assay in 24- or 96-well plates. J. Virol. Methods 33:191-198. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
