

Elastic coupling of integral membrane protein stability to lipid bilayer forces
- PMID: 14990786
- PMCID: PMC384696
- DOI: 10.1073/pnas.0400358101
Elastic coupling of integral membrane protein stability to lipid bilayer forces
Abstract
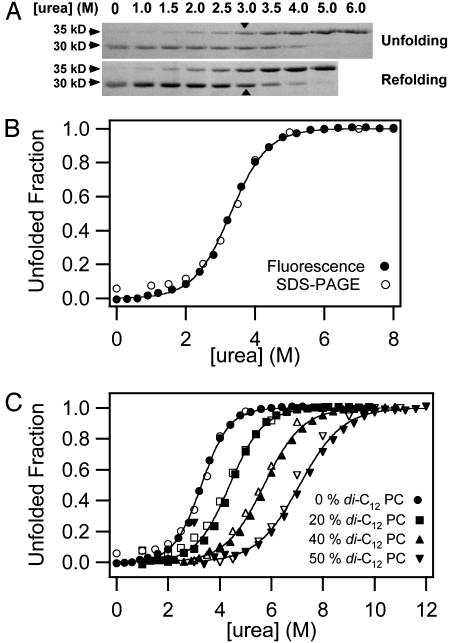
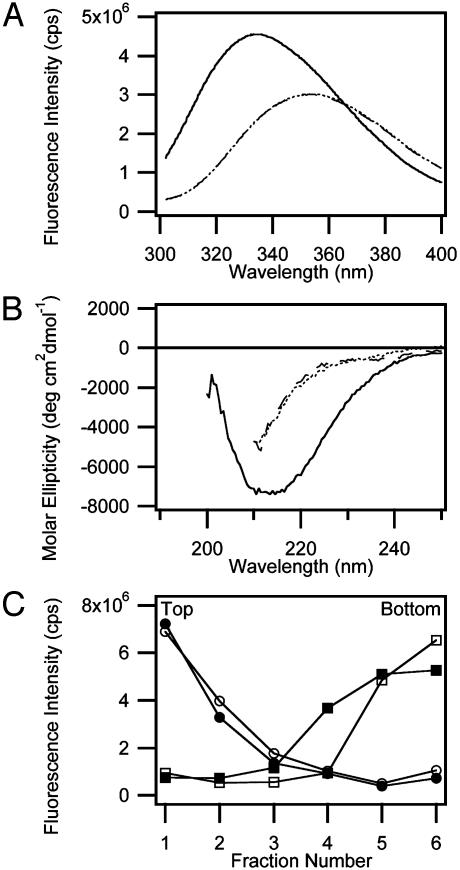
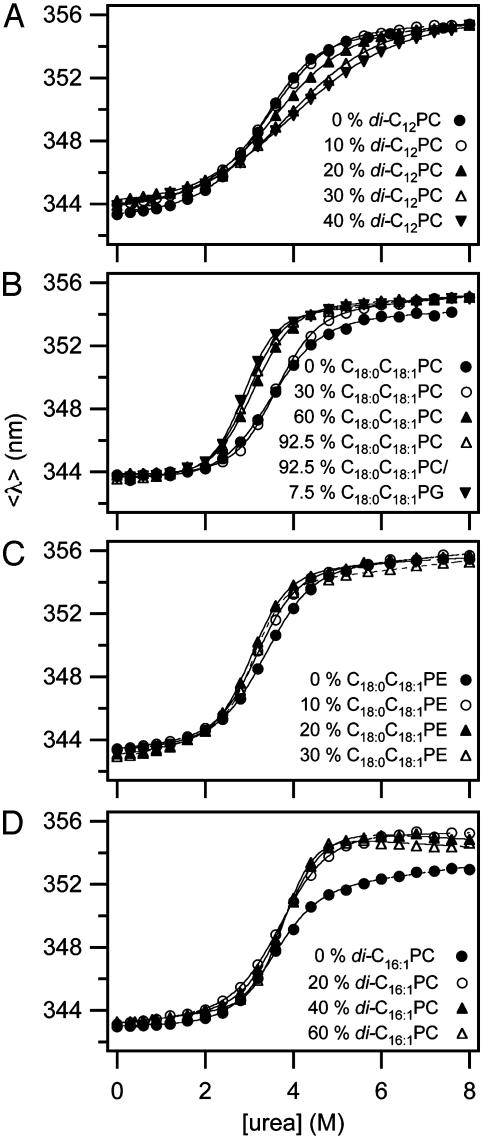
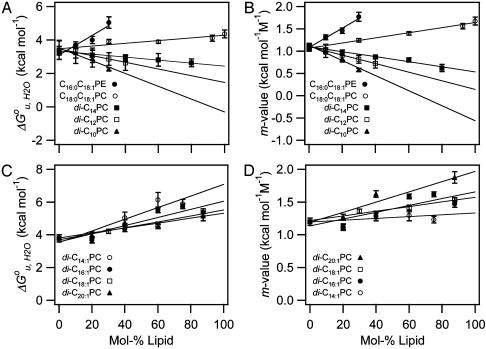
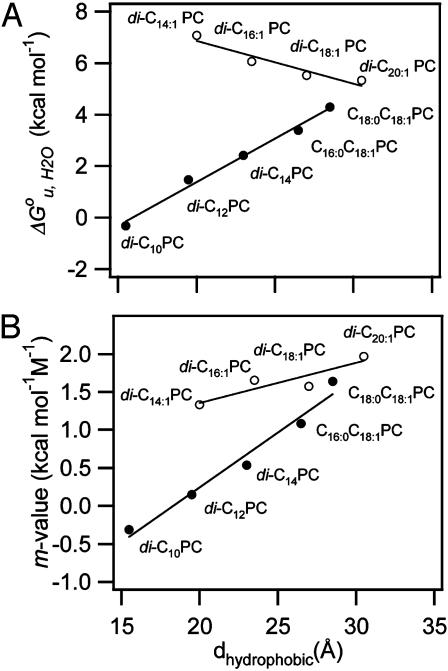
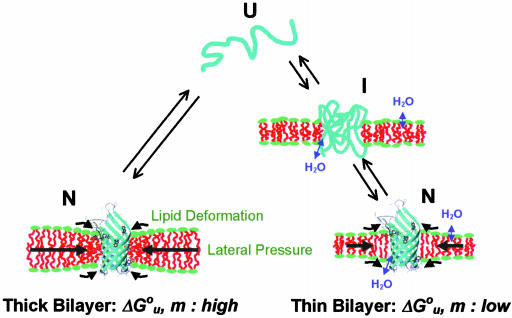
It has been traditionally difficult to measure the thermodynamic stability of membrane proteins because fully reversible protocols for complete folding these proteins were not available. Knowledge of the thermodynamic stability of membrane proteins is desirable not only from a fundamental theoretical standpoint, but is also of enormous practical interest for the rational design of membrane proteins and for optimizing conditions for their structure determination by crystallography or NMR. Here, we describe the design of a fully reversible system to study equilibrium folding of the outer membrane protein A from Escherichia coli in lipid bilayers. Folding is shown to be two-state under appropriate conditions permitting data analysis with a classical folding model developed for soluble proteins. The resulting free energy and m value, i.e., a measure of cooperativity, of unfolding are DeltaG(u,H2O)(o)=3.4 kcal/mol and m = 1.1 kcal/mol M(-1), respectively, in a reference bilayer composed of palmitoyl-oleoyl-phosphatidylcholine (C(16:0)C(18:1)PC) and palmitoyloleoyl-phosphatidylglycerol (C(16:0)C(18:1)PG). These values are strong functions of the lipid bilayer environment. By systematic variation of lipid headgroup and chain composition, we show that elastic bilayer forces such as curvature stress and hydrophobic mismatch modulate the free energy and cooperativity of folding of this and perhaps many other membrane proteins.
Figures
Comment in
-
Membrane proteins: a new method enters the fold.Proc Natl Acad Sci U S A. 2004 Mar 23;101(12):3995-6. doi: 10.1073/pnas.0400671101. Epub 2004 Mar 15. Proc Natl Acad Sci U S A. 2004. PMID: 15024105 Free PMC article. No abstract available.
References
-
- Martinac, B., Adler, J. & Kung, C. (1990) Nature 348, 261–263. - PubMed
-
- Perozo, E., Cortes, D. M., Sompornpisut, P., Kloda, A. & Martinac, B. (2002) Nature 418, 942–948. - PubMed
-
- Brown, M. F. (1994) Chem. Phys. Lipids 73, 159–180. - PubMed
-
- Navarro, J., Toivio-Kinnucan, M. & Racker, E. (1984) Biochemistry 23, 130–135. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
