

Sox5 and Sox6 are needed to develop and maintain source, columnar, and hypertrophic chondrocytes in the cartilage growth plate
- PMID: 14993235
- PMCID: PMC2172159
- DOI: 10.1083/jcb.200312045
Sox5 and Sox6 are needed to develop and maintain source, columnar, and hypertrophic chondrocytes in the cartilage growth plate
Abstract
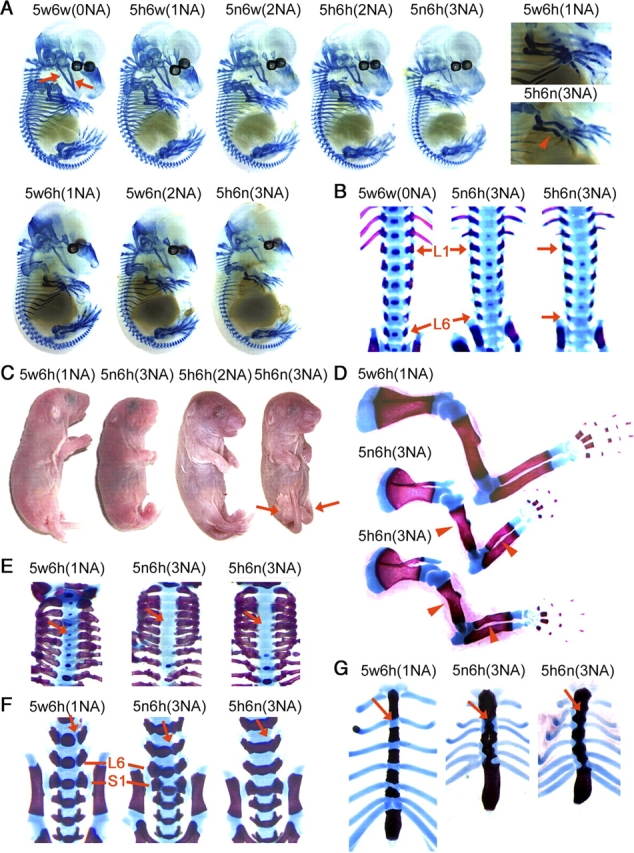
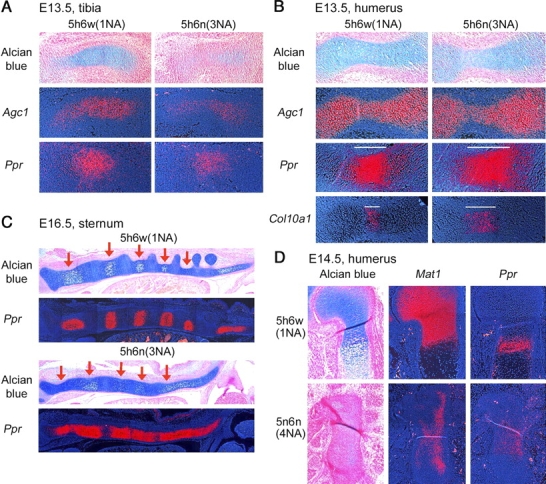
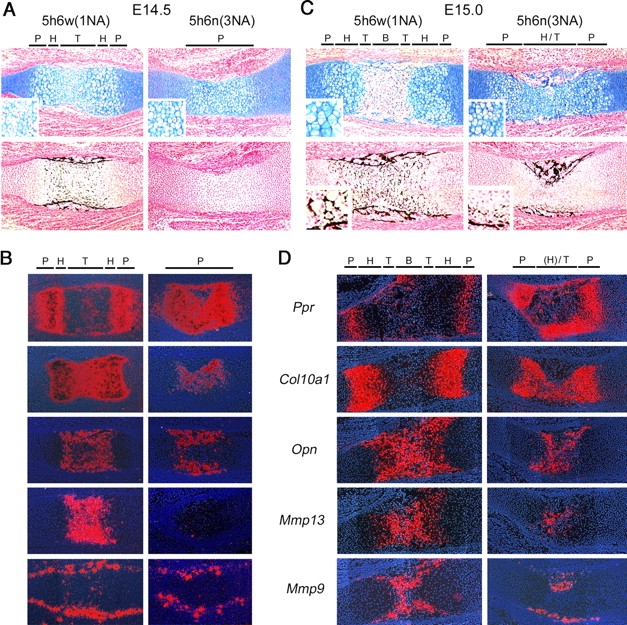
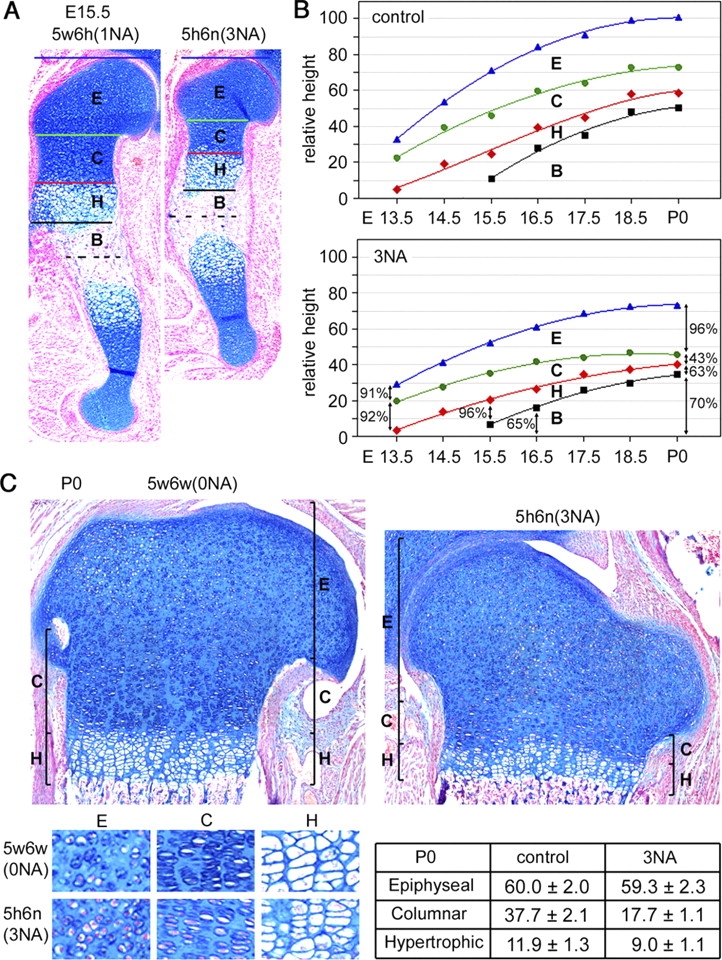
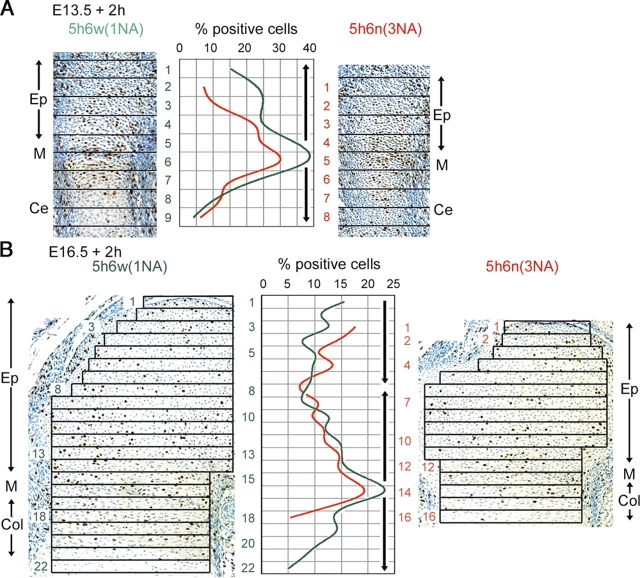
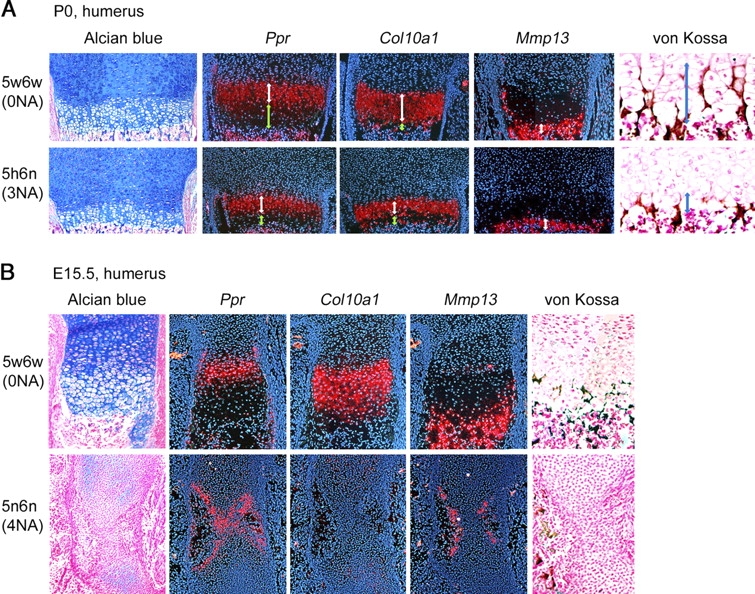
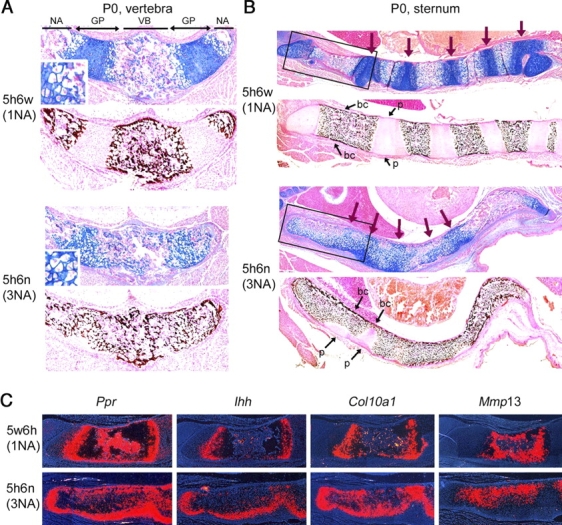
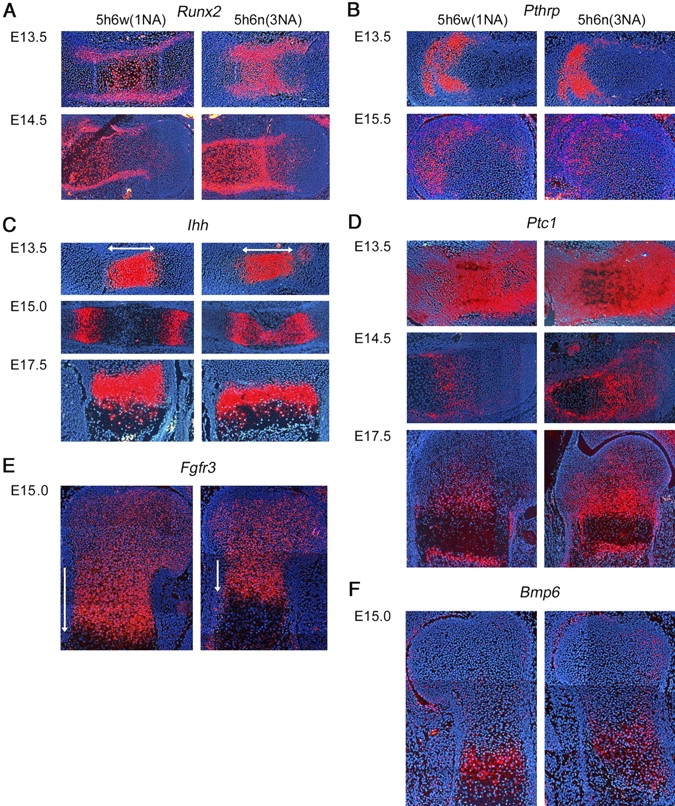
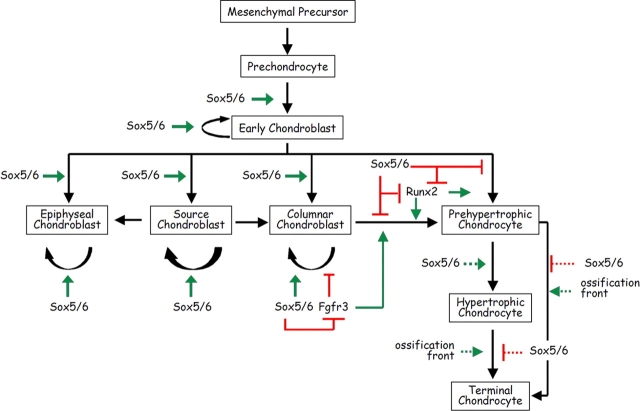
Sox5 and Sox6 encode Sry-related transcription factors that redundantly promote early chondroblast differentiation. Using mouse embryos with three or four null alleles of Sox5 and Sox6, we show that they are also essential and redundant in major steps of growth plate chondrocyte differentiation. Sox5 and Sox6 promote the development of a highly proliferating pool of chondroblasts between the epiphyses and metaphyses of future long bones. This pool is the likely cellular source of growth plates. Sox5 and Sox6 permit formation of growth plate columnar zones by keeping chondroblasts proliferating and by delaying chondrocyte prehypertrophy. They allow induction of chondrocyte hypertrophy and permit formation of prehypertrophic and hypertrophic zones by delaying chondrocyte terminal differentiation induced by ossification fronts. They act, at least in part, by down-regulating Ihh signaling, Fgfr3, and Runx2 and by up-regulating Bmp6. In conclusion, Sox5 and Sox6 are needed for the establishment of multilayered growth plates, and thereby for proper and timely development of endochondral bones.
Figures
References
-
- Abad, V., J.L. Meyers, M. Weise, R.I. Gafni, K.M. Barnes, O. Nilsson, J.D. Bacher, and J. Baron. 2002. The role of the resting zone in growth plate chondrogenesis. Endocrinology. 143:1851–1857. - PubMed
-
- Baur, S.T., J.J. Mai, and S.M. Dymecki. 2000. Combinatorial signaling through BMP receptor IB and GDF5: shaping of the distal mouse limb and the genetics of distal limb diversity. Development. 127:605–619. - PubMed
-
- Bi, W., J.M. Deng, Z. Zhang, R.R. Behringer, and B. de Crombrugghe. 1999. Sox9 is required for cartilage formation. Nat. Genet. 22:85–89. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
