

The herpes simplex virus 1 UL41 gene-dependent destabilization of cellular RNAs is selective and may be sequence-specific
- PMID: 14993598
- PMCID: PMC373509
- DOI: 10.1073/pnas.0400354101
The herpes simplex virus 1 UL41 gene-dependent destabilization of cellular RNAs is selective and may be sequence-specific
Abstract
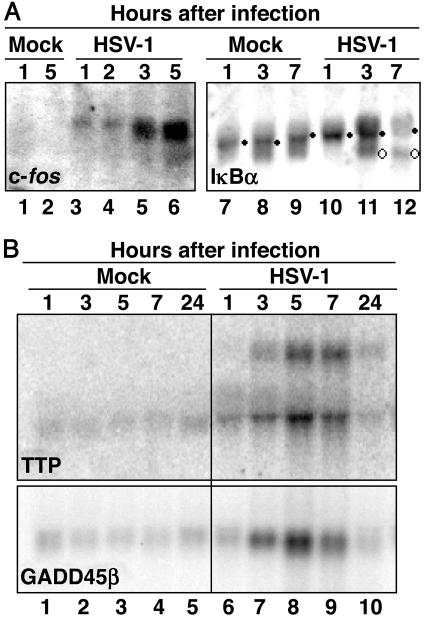
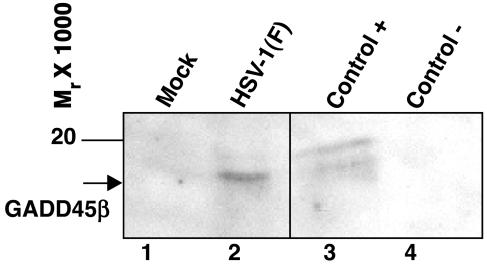
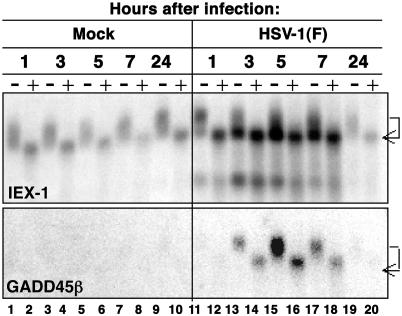
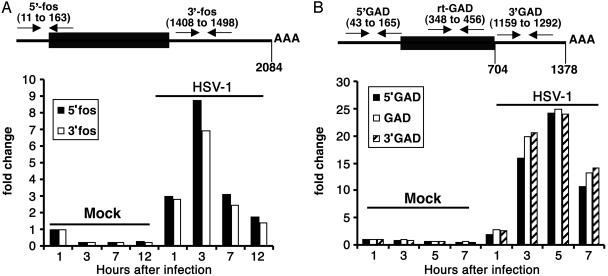
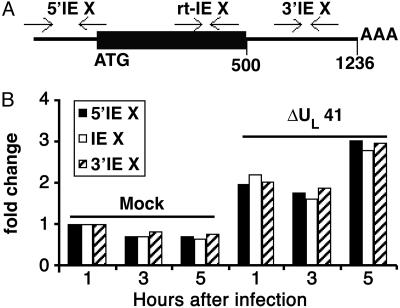
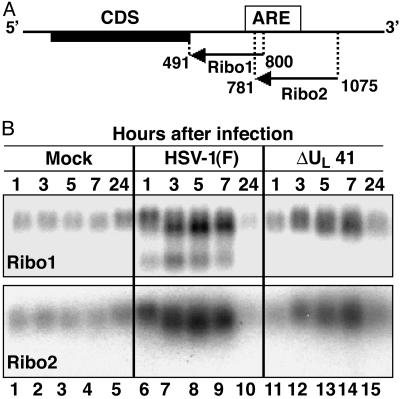
In cells infected with herpes simplex virus 1, the RNA encoded by the stress-inducible immediate early response gene IEX-1 was up-regulated immediately after infection. However, the accumulated RNA was degraded 3'-5', and the protein was detectable only at very early times after infection. The degradation was dependent on the U(L)41 gene encoding the virion host shutoff (vhs) protein and resulted in the accumulation of truncated RNA containing the 5'-end portion of the transcript. IEX-1 contains an AU-rich element (ARE) in its 3'-untranslated domains known to regulate negatively the RNA lifespan. To examine the role of ARE in signaling the degradation, we compared the stability of several RNAs up-regulated during infection to WT virus. These were ARE-containing RNAs encoding IEX-1, c-fos, and IkappaBalpha and the non-ARE-containing RNAs GADD45beta and tristetraprolin. We report that the ARE-containing RNAs exemplified by IEX-1 RNA are deadenylated and cleaved in the ARE within the 3' UTR in a U(L)41-dependent manner. In contrast, Northern blot hybridizations and analyses of poly(A) tails revealed no evidence of degradation of GADD45beta RNA. GADD45beta protein was detected in WT virus-infected cells. These results indicate that the degradation of RNAs and the mechanism by which cellular RNAs are degraded are selective and may be sequence specific. The persistence of partially degraded ARE-containing RNAs may reflect specific targeting of the vhs proteins to the ARE and the modification of the RNA degradation machinery of the cell induced by the presence of the vhs protein.
Figures
References
-
- Karr, B. M. & Read, G. S. (1999) Virology 264, 195-204. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
