

A variable dinucleotide repeat in the CFTR gene contributes to phenotype diversity by forming RNA secondary structures that alter splicing
- PMID: 14993601
- PMCID: PMC373492
- DOI: 10.1073/pnas.0400182101
A variable dinucleotide repeat in the CFTR gene contributes to phenotype diversity by forming RNA secondary structures that alter splicing
Abstract
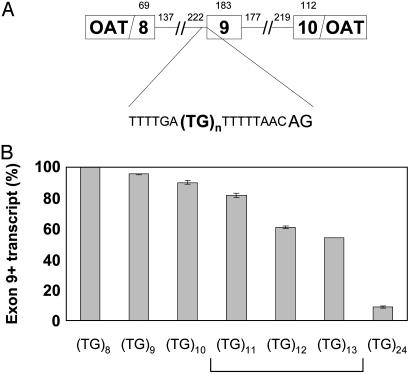
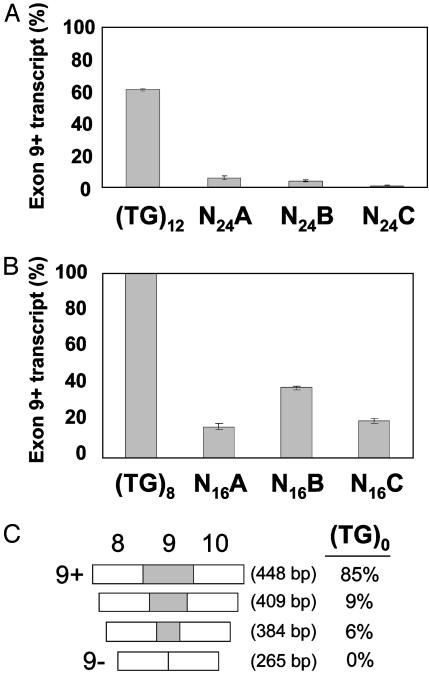
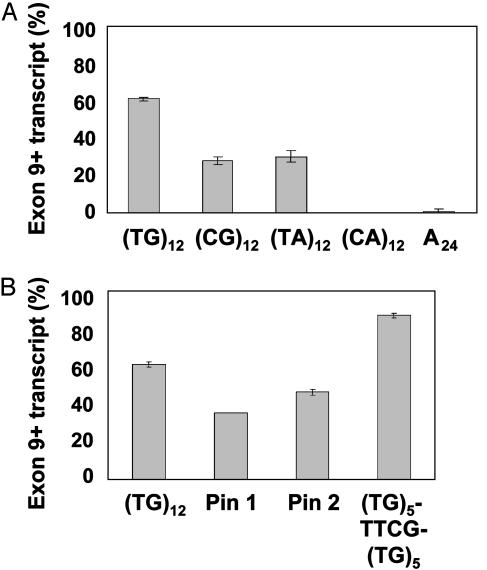
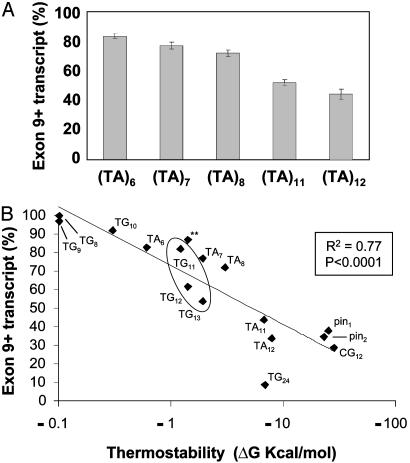
Dinucleotide repeats are ubiquitous features of eukaryotic genomes that are not generally considered to have functional roles in gene expression. However, the highly variable nature of dinucleotide repeats makes them particularly interesting candidates for modifiers of RNA splicing when they are found near splicing signals. An example of a variable dinucleotide repeat that affects splicing is a TG repeat located in the splice acceptor of exon 9 of the cystic fibrosis transmembrane conductance regulator (CFTR) gene. Higher repeat numbers result in reduced exon 9 splicing efficiency and, in some instances, the reduction in full-length transcript is sufficient to cause male infertility due to congenital bilateral absence of the vas deferens or nonclassic cystic fibrosis. Using a CFTR minigene system, we studied TG tract variation and observed the same correlation between dinucleotide repeat number and exon 9 splicing efficiency seen in vivo. Replacement of the TG dinucleotide tract in the minigene with random sequence abolished splicing of exon 9. Replacements of the TG tract with sequences that can self-base-pair suggested that the formation of an RNA secondary structure was associated with efficient splicing. However, splicing efficiency was inversely correlated with the predicted thermodynamic stability of such structures, demonstrating that intermediate stability was optimal. Finally, substitution with TA repeats of differing length confirmed that stability of the RNA secondary structure, not sequence content, correlated with splicing efficiency. Taken together, these data indicate that dinucleotide repeats can form secondary structures that have variable effects on RNA splicing efficiency and clinical phenotype.
Figures
References
-
- Gabellini, N. (2001) Eur. J. Biochem. 268, 1076-1083. - PubMed
-
- Hui, J., Stangl, K., Lane, W. S. & Bindereif, A. (2003) Nat. Struct. Biol. 10, 33-37. - PubMed
-
- Stangl, K., Cascorbi, I., Laule, M., Klein, T., Stangl, V., Rost, S., Wernecke, K. D., Felix, S. B., Bindereif, A., Baumann, G., et al. (2000) Pharmacogenetics 10, 133-140. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
