

The Legionella pneumophila PilT homologue DotB exhibits ATPase activity that is critical for intracellular growth
- PMID: 14996796
- PMCID: PMC355965
- DOI: 10.1128/JB.186.6.1658-1666.2004
The Legionella pneumophila PilT homologue DotB exhibits ATPase activity that is critical for intracellular growth
Abstract
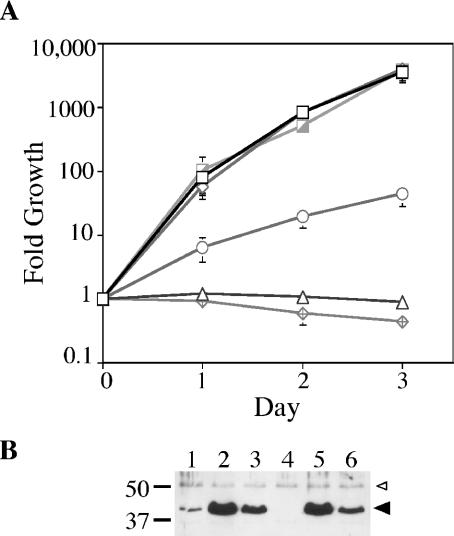
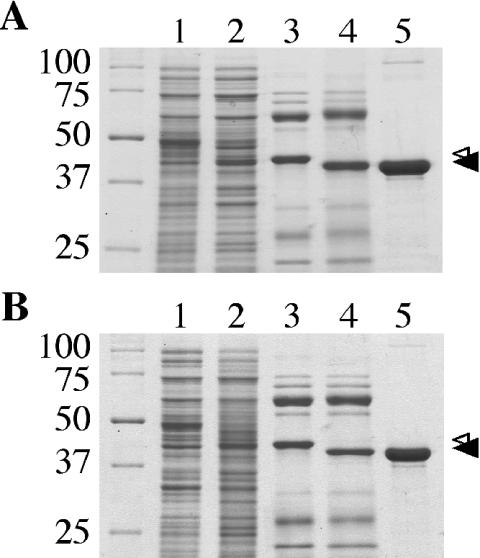
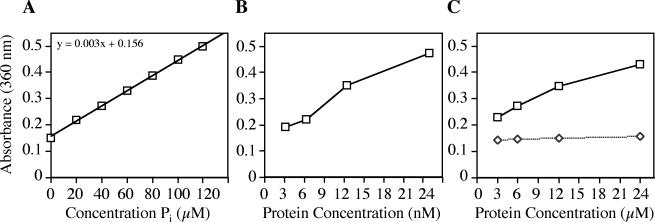
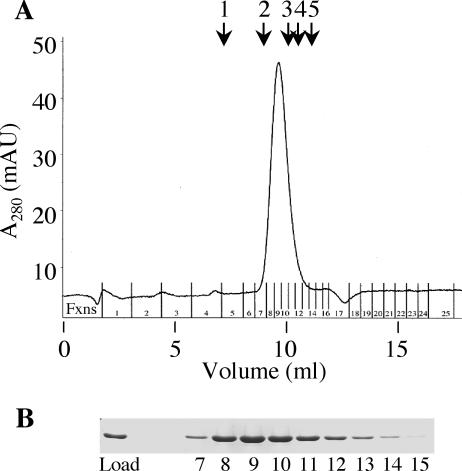
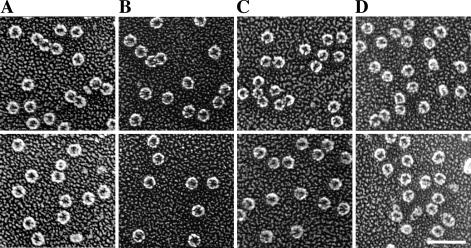
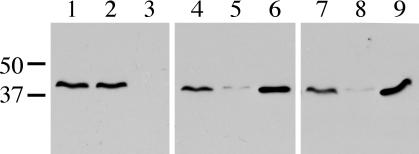
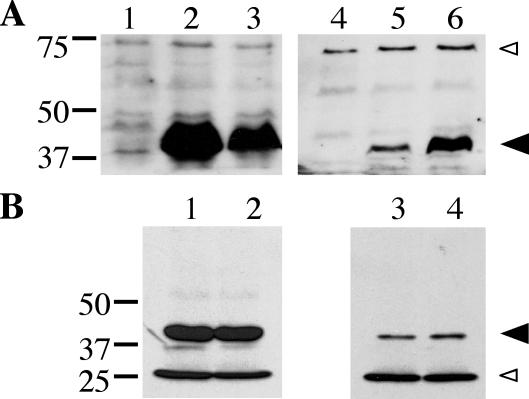
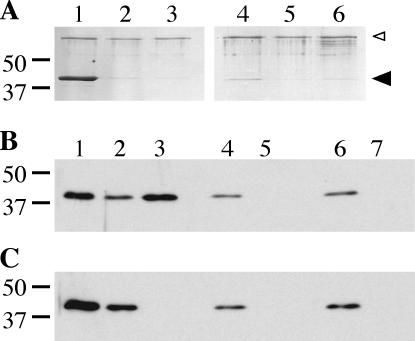
The ability of Legionella pneumophila to grow and cause disease in the host is completely dependent on a type IV secretion system known as the Dot/Icm complex. This membrane-spanning apparatus translocates effector molecules into host cells in a process that is poorly understood but that is known to require the putative ATPase DotB. One possible role for DotB is suggested by its similarity to the PilT family of proteins, which mediate pilus retraction. To better understand the molecular behavior of DotB, we have purified the protein and shown that it forms stable homohexameric rings and hydrolyzes ATP with a specific activity of 6.4 nmol of ATP/min/mg of protein. ATPase activity is critical to the function of DotB, as alteration of the conserved Walker box lysine residue resulted in a mutant protein, DotB K162Q, which failed to bind or hydrolyze ATP and which could not complement a DeltadotB strain for intracellular growth in macrophages. Consistent with the ability of DotB to interact with itself, the dotBK162Q allele exhibited transdominance over wild-type dotB, providing the first example of such a mutation in L. pneumophila. Finally, the DotB K162Q mutant protein had a significantly enhanced membrane localization in L. pneumophila compared to wild-type DotB, suggesting a relationship between nucleotide binding and membrane association. These results are consistent with a model in which DotB cycles between the cytoplasm and the Dot/Icm complex at the membrane, where it hydrolyzes nucleotides to provide energy to the complex.
Figures
References
-
- Berger, K. H., and R. R. Isberg. 1993. Two distinct defects in intracellular growth complemented by a single genetic locus in Legionella pneumophila. Mol. Microbiol. 7:7-19. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
