

The pKO2 linear plasmid prophage of Klebsiella oxytoca
- PMID: 14996813
- PMCID: PMC355964
- DOI: 10.1128/JB.186.6.1818-1832.2004
The pKO2 linear plasmid prophage of Klebsiella oxytoca
Abstract
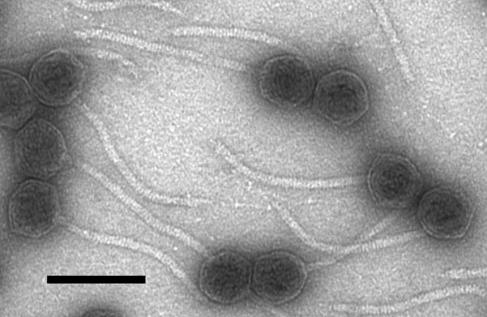
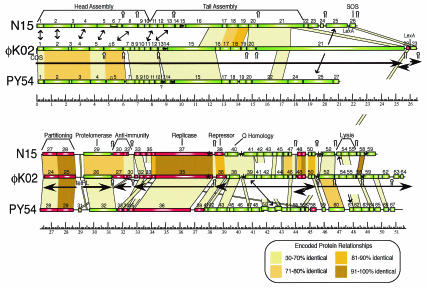
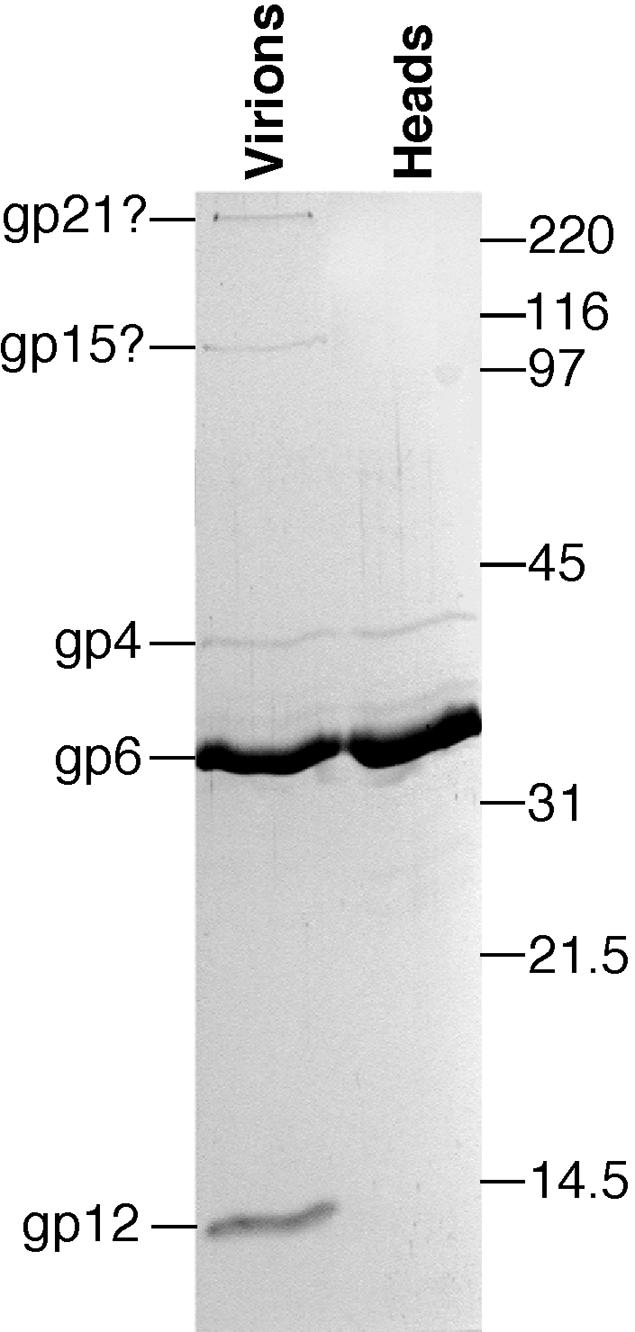
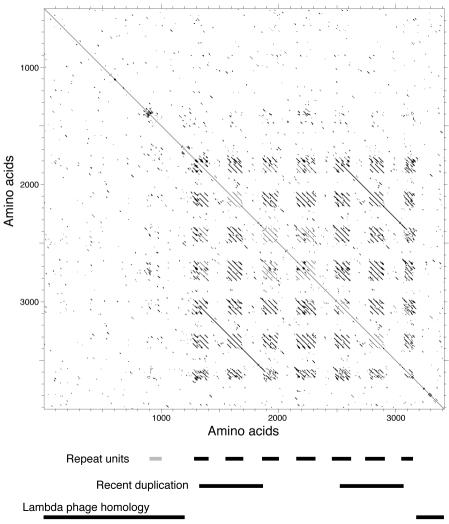
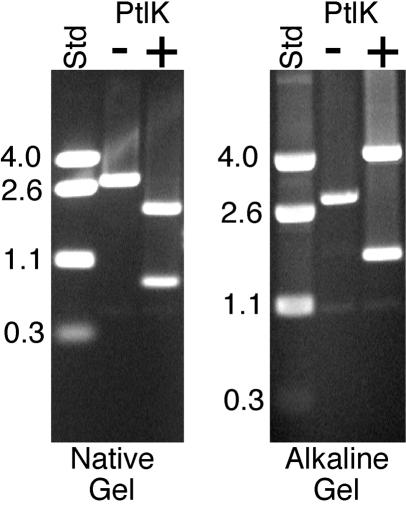
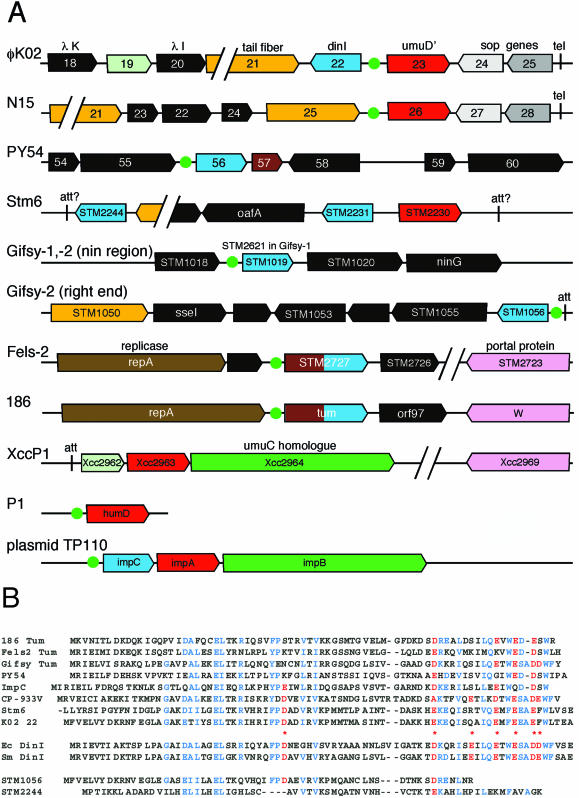
Temperate bacteriophages with plasmid prophages are uncommon in nature, and of these only phages N15 and PY54 are known to have a linear plasmid prophage with closed hairpin telomeres. We report here the complete nucleotide sequence of the 51,601-bp Klebsiella oxytoca linear plasmid pKO2, and we demonstrate experimentally that it is also a prophage. We call this bacteriophage phiKO2. An analysis of the 64 predicted phiKO2 genes indicate that it is a fairly close relative of phage N15; they share a mosaic relationship that is typical of different members of double-stranded DNA tailed-phage groups. Although the head, tail shaft, and lysis genes are not recognizably homologous between these phages, other genes such as the plasmid partitioning, replicase, prophage repressor, and protelomerase genes (and their putative targets) are so similar that we predict that they must have nearly identical DNA binding specificities. The phiKO2 virion is unusual in that its phage lambda-like tails have an exceptionally long (3,433 amino acids) central tip tail fiber protein. The phiKO2 genome also carries putative homologues of bacterial dinI and umuD genes, both of which are involved in the host SOS response. We show that these divergently transcribed genes are regulated by LexA protein binding to a single target site that overlaps both promoters.
Figures

Similar articles
-
N15: the linear phage-plasmid.Plasmid. 2011 Mar;65(2):102-9. doi: 10.1016/j.plasmid.2010.12.004. Epub 2010 Dec 23. Plasmid. 2011. PMID: 21185326 Review.
-
Interplay between the temperate phages PY54 and N15, linear plasmid prophages with covalently closed ends.J Bacteriol. 2007 Nov;189(22):8366-70. doi: 10.1128/JB.01066-07. Epub 2007 Sep 7. J Bacteriol. 2007. PMID: 17827299 Free PMC article.
-
Sequence analysis of the genome of the temperate Yersinia enterocolitica phage PY54.J Mol Biol. 2003 Aug 15;331(3):605-22. doi: 10.1016/s0022-2836(03)00763-0. J Mol Biol. 2003. PMID: 12899832
-
Genomic sequence and analysis of the atypical temperate bacteriophage N15.J Mol Biol. 2000 May 26;299(1):53-73. doi: 10.1006/jmbi.2000.3731. J Mol Biol. 2000. PMID: 10860722
-
Replication and Maintenance of Linear Phage-Plasmid N15.Microbiol Spectr. 2015 Feb;3(1):PLAS-0032-2014. doi: 10.1128/microbiolspec.PLAS-0032-2014. Microbiol Spectr. 2015. PMID: 26104561 Review.
Cited by
-
Identification, Characterization, and Genomic Analysis of Novel Serratia Temperate Phages from a Gold Mine.Int J Mol Sci. 2020 Sep 13;21(18):6709. doi: 10.3390/ijms21186709. Int J Mol Sci. 2020. PMID: 32933193 Free PMC article.
-
Error-prone DNA repair system in enteroaggregative Escherichia coli identified by subtractive hybridization.J Bacteriol. 2007 May;189(10):3793-803. doi: 10.1128/JB.01764-06. Epub 2007 Mar 9. J Bacteriol. 2007. PMID: 17351038 Free PMC article.
-
The repA gene of the linear Yersinia enterocolitica prophage PY54 functions as a circular minimal replicon in Escherichia coli.J Bacteriol. 2005 May;187(10):3445-54. doi: 10.1128/JB.187.10.3445-3454.2005. J Bacteriol. 2005. PMID: 15866931 Free PMC article.
-
Characteristics of phage-plasmids and their impact on microbial communities.Essays Biochem. 2024 Dec 17;68(5):583-592. doi: 10.1042/EBC20240014. Essays Biochem. 2024. PMID: 39611587 Free PMC article. Review.
-
Linear chromosome-generating system of Agrobacterium tumefaciens C58: protelomerase generates and protects hairpin ends.J Biol Chem. 2012 Jul 20;287(30):25551-63. doi: 10.1074/jbc.M112.369488. Epub 2012 May 10. J Biol Chem. 2012. PMID: 22582388 Free PMC article.
References
-
- Allison, G. E., D. C. Angeles, P. Huan, and N. K. Verma. 2003. Morphology of temperate bacteriophage SfV and characterisation of the DNA packaging and capsid genes: the structural genes evolved from two different phage families. Virology 308:114-127. - PubMed
-
- Berkmen, M., and M. J. Benedik. 2002. Multi-copy repression of Serratia marcescens nuclease expression by dinI. Curr. Microbiol. 44:44-48. - PubMed
-
- Biere, A. L., M. Citron, and H. Schuster. 1992. Transcriptional control via translational repression by c4 antisense RNA of bacteriophages P1 and P7. Genes Dev. 6:2409-2416. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
