

Identification and characterization of two novel proteins affecting fission yeast gamma-tubulin complex function
- PMID: 15004232
- PMCID: PMC404023
- DOI: 10.1091/mbc.e03-10-0728
Identification and characterization of two novel proteins affecting fission yeast gamma-tubulin complex function
Abstract
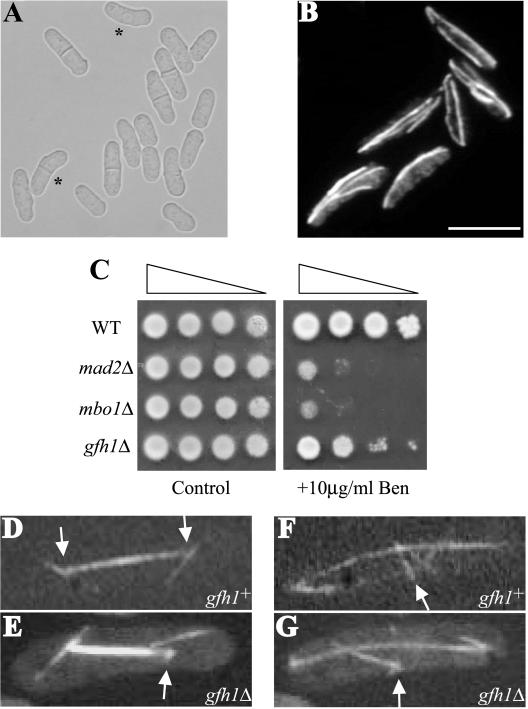
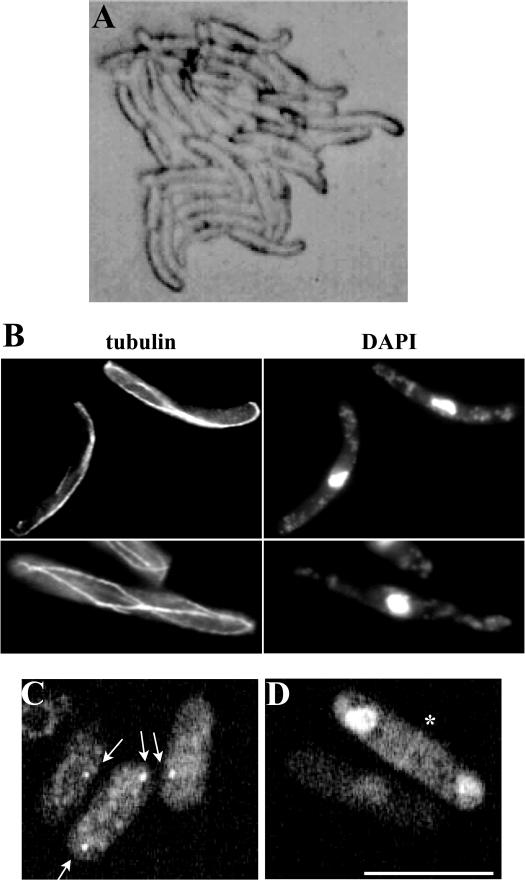
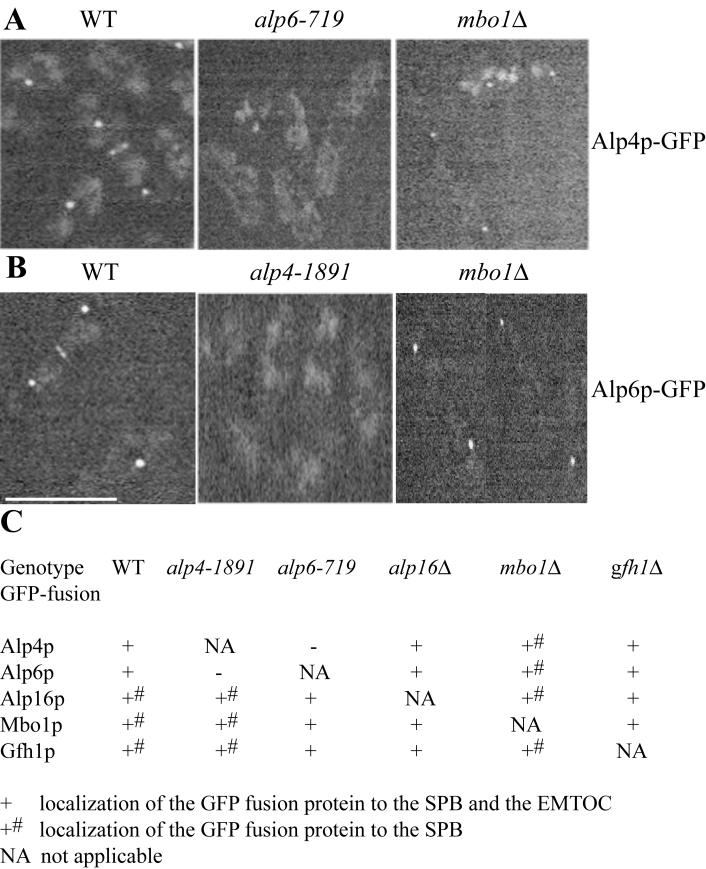
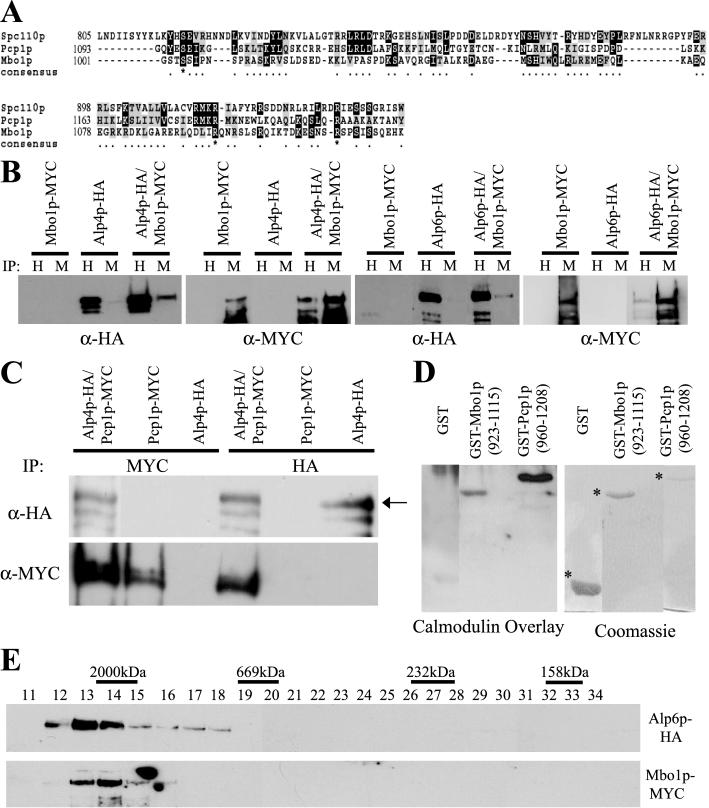
The gamma-tubulin complex, via its ability to organize microtubules, is critical for accurate chromosome segregation and cytokinesis in the fission yeast, Schizosaccharomyces pombe. To better understand its roles, we have purified the S. pombe gamma-tubulin complex. Mass spectrometric analyses of the purified complex revealed known components and identified two novel proteins (i.e., Mbo1p and Gfh1p) with homology to gamma-tubulin-associated proteins from other organisms. We show that both Mbo1p and Gfh1p localize to microtubule organizing centers. Although cells deleted for either mbo1(+) or gfh1(+) are viable, they exhibit a number of defects associated with altered microtubule function such as defects in cell polarity, nuclear positioning, spindle orientation, and cleavage site specification. In addition, mbo1Delta and gfh1Delta cells exhibit defects in astral microtubule formation and anchoring, suggesting that these proteins have specific roles in astral microtubule function. This study expands the known roles of gamma-tubulin complex components in organizing different types of microtubule structures in S. pombe.
Figures



Similar articles
-
Mto2p, a novel fission yeast protein required for cytoplasmic microtubule organization and anchoring of the cytokinetic actin ring.Mol Biol Cell. 2005 Jun;16(6):3052-63. doi: 10.1091/mbc.e04-12-1043. Epub 2005 Mar 30. Mol Biol Cell. 2005. PMID: 15800064 Free PMC article.
-
Fission yeast mto2p regulates microtubule nucleation by the centrosomin-related protein mto1p.Mol Biol Cell. 2005 Jun;16(6):3040-51. doi: 10.1091/mbc.e04-11-1003. Epub 2005 Jan 19. Mol Biol Cell. 2005. PMID: 15659644 Free PMC article.
-
Noncore components of the fission yeast gamma-tubulin complex.Mol Biol Cell. 2006 Dec;17(12):5075-93. doi: 10.1091/mbc.e05-11-1009. Epub 2006 Oct 4. Mol Biol Cell. 2006. PMID: 17021256 Free PMC article.
-
Big Lessons from Little Yeast: Budding and Fission Yeast Centrosome Structure, Duplication, and Function.Annu Rev Genet. 2017 Nov 27;51:361-383. doi: 10.1146/annurev-genet-120116-024733. Epub 2017 Sep 15. Annu Rev Genet. 2017. PMID: 28934593 Review.
-
[Is spindle formation in fission yeast specific to the species?: from the viewpoint of nuclear transport and spindle pole body].Tanpakushitsu Kakusan Koso. 2008 Mar;53(3):197-206. Tanpakushitsu Kakusan Koso. 2008. PMID: 18326292 Review. Japanese. No abstract available.
Cited by
-
Mto2p, a novel fission yeast protein required for cytoplasmic microtubule organization and anchoring of the cytokinetic actin ring.Mol Biol Cell. 2005 Jun;16(6):3052-63. doi: 10.1091/mbc.e04-12-1043. Epub 2005 Mar 30. Mol Biol Cell. 2005. PMID: 15800064 Free PMC article.
-
Drosophila melanogaster gamma-TuRC is dispensable for targeting gamma-tubulin to the centrosome and microtubule nucleation.J Cell Biol. 2006 Feb 13;172(4):517-28. doi: 10.1083/jcb.200511071. J Cell Biol. 2006. PMID: 16476773 Free PMC article.
-
CDK5RAP2 stimulates microtubule nucleation by the gamma-tubulin ring complex.J Cell Biol. 2010 Dec 13;191(6):1089-95. doi: 10.1083/jcb.201007030. Epub 2010 Dec 6. J Cell Biol. 2010. PMID: 21135143 Free PMC article.
-
Activation of the γ-tubulin complex by the Mto1/2 complex.Curr Biol. 2014 Apr 14;24(8):896-903. doi: 10.1016/j.cub.2014.03.006. Epub 2014 Apr 3. Curr Biol. 2014. PMID: 24704079 Free PMC article.
-
Latrunculin A delays anaphase onset in fission yeast by disrupting an Ase1-independent pathway controlling mitotic spindle stability.Mol Biol Cell. 2008 Sep;19(9):3713-23. doi: 10.1091/mbc.e08-02-0164. Epub 2008 Jun 18. Mol Biol Cell. 2008. PMID: 18562692 Free PMC article.
References
-
- Bahler, J., Wu, J.Q., Longtine, M.S., Shah, N.G., McKenzie, A., 3rd, Steever, A.B., Wach, A., Philippsen, P., and Pringle, J.R. (1998). Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast 14, 943–951. - PubMed
-
- Basi, G., Schmid, E., and Maundrell, K. (1993). TATA box mutations in the Schizosaccharomyces pombe nmt1 promoter affect transcription efficiency but not the transcription start point or thiamine repressibility. Gene 123, 131–136. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
