

The role and regulation of the preRC component Cdc6 in the initiation of premeiotic DNA replication
- PMID: 15004237
- PMCID: PMC404018
- DOI: 10.1091/mbc.e03-08-0617
The role and regulation of the preRC component Cdc6 in the initiation of premeiotic DNA replication
Abstract
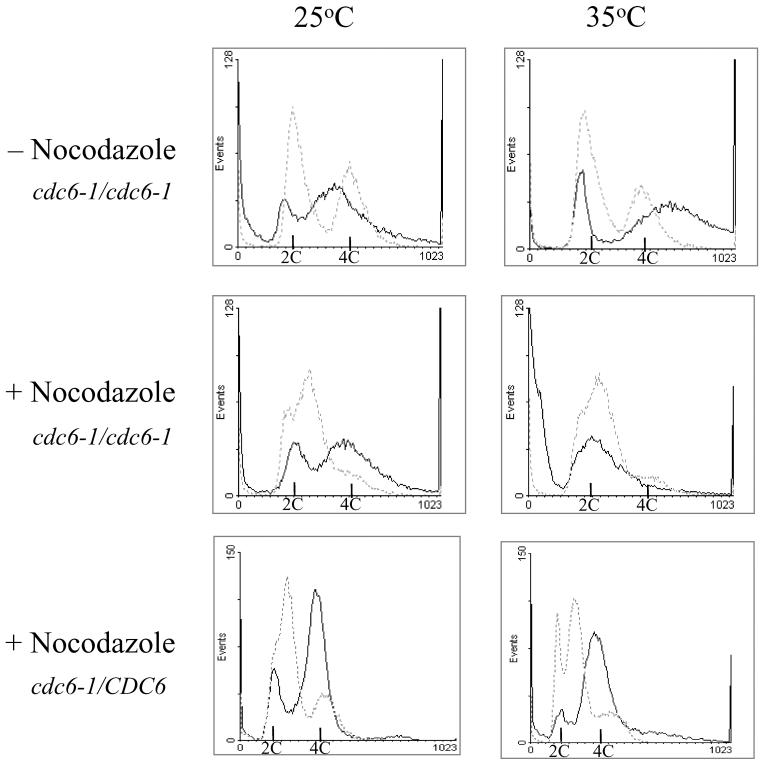
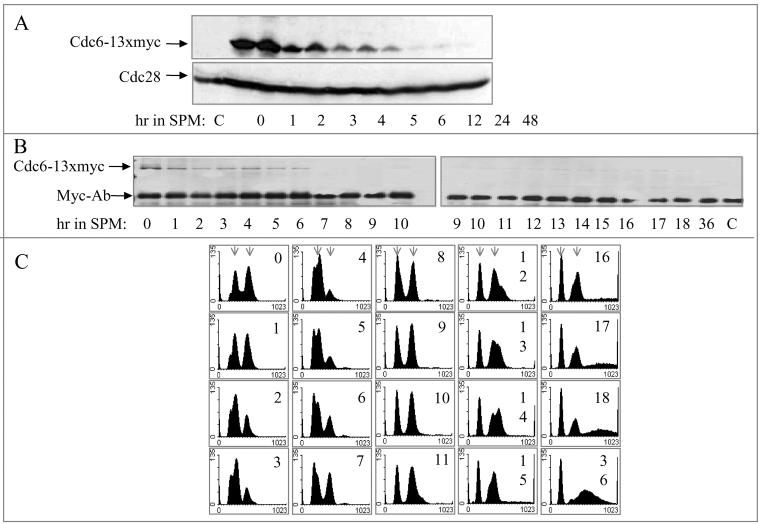
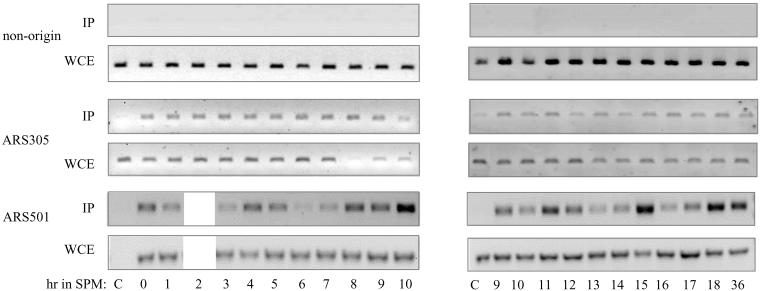
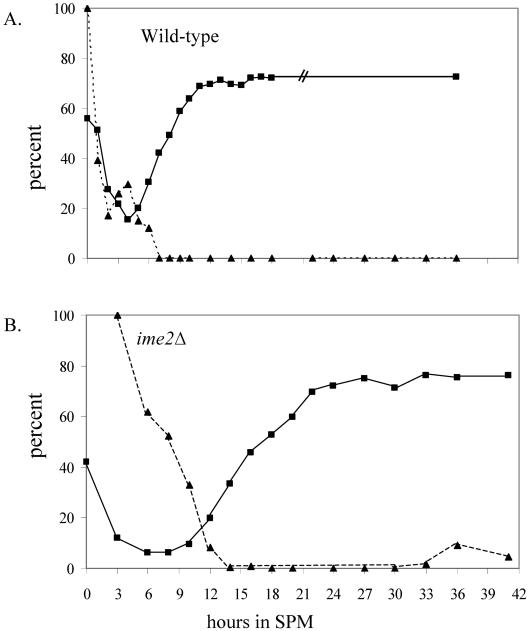
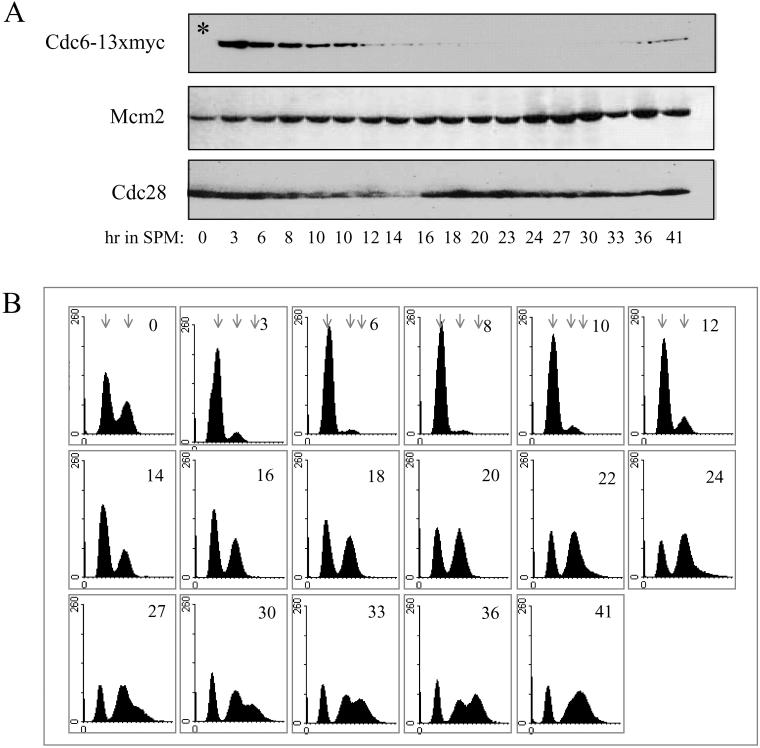
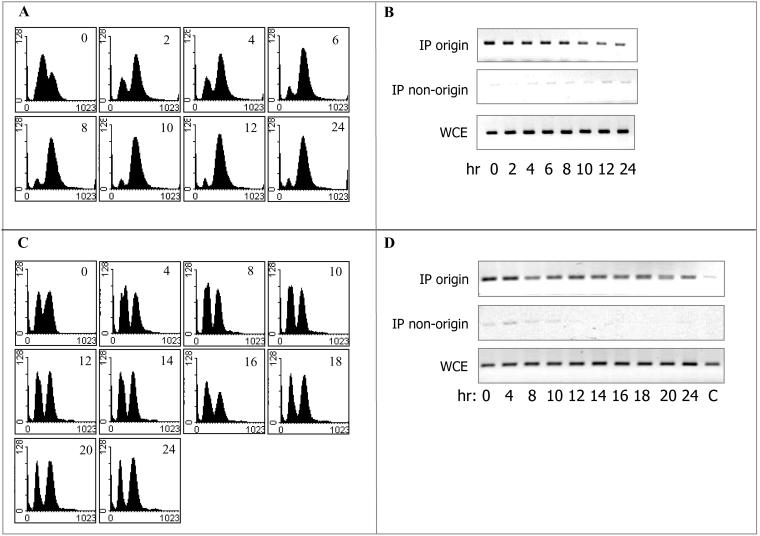
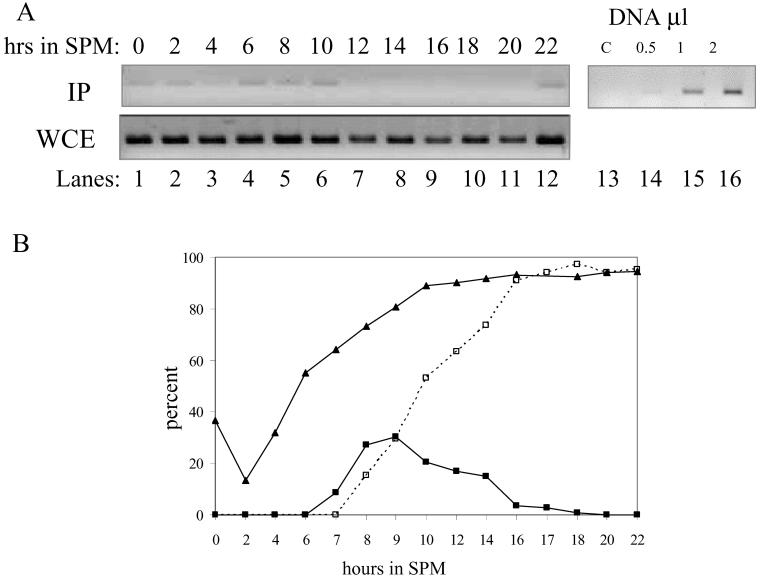
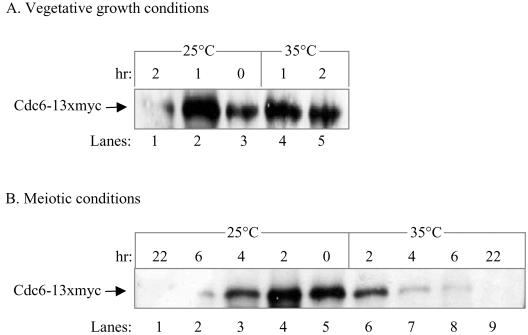
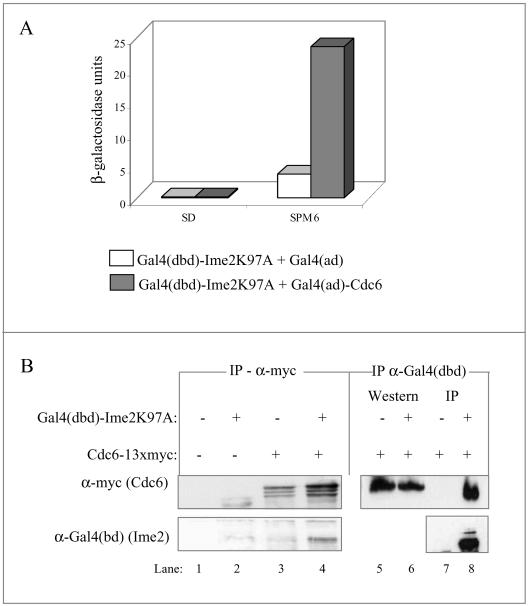
In all eukaryotes, the initiation of DNA replication is regulated by the ordered assembly of DNA/protein complexes on origins of DNA replication. In this report, we examine the role of Cdc6, a component of the prereplication complex, in the initiation of premeiotic DNA replication in budding yeast. We show that in the meiotic cycle, Cdc6 is required for DNA synthesis and sporulation. Moreover, similarly to the regulation in the mitotic cell cycle, Cdc6 is specifically degraded upon entry into the meiotic S phase. By contrast, chromatin-immunoprecipitation analysis reveals that the origin-bound Cdc6 is stable throughout the meiotic cycle. Preliminary evidence suggests that this protection reflects a change in chromatin structure that occurs in meiosis. Using the cdc28-degron allele, we show that depletion of Cdc28 leads to stabilization of Cdc6 in the mitotic cycle, but not in the meiotic cycle. We show physical association between Cdc6 and the meiosis-specific hCDK2 homolog Ime2. These results suggest that under meiotic conditions, Ime2, rather than Cdc28, regulates the stability of Cdc6. Chromatin-immunoprecipitation analysis reveals that similarly to the mitotic cell cycle, Mcm2 binds origins in G1 and meiotic S phases, and at the end of the second meiotic division, it is gradually removed from chromatin.
Figures
Similar articles
-
Cdc28 and Ime2 possess redundant functions in promoting entry into premeiotic DNA replication in Saccharomyces cerevisiae.Genetics. 2001 Dec;159(4):1547-58. doi: 10.1093/genetics/159.4.1547. Genetics. 2001. PMID: 11779796 Free PMC article.
-
Control of landmark events in meiosis by the CDK Cdc28 and the meiosis-specific kinase Ime2.Genes Dev. 2003 Jun 15;17(12):1524-39. doi: 10.1101/gad.1101503. Epub 2003 Jun 3. Genes Dev. 2003. PMID: 12783856 Free PMC article.
-
Clb/Cdc28 kinases promote nuclear export of the replication initiator proteins Mcm2-7.Curr Biol. 2000 Feb 24;10(4):195-205. doi: 10.1016/s0960-9822(00)00337-7. Curr Biol. 2000. PMID: 10704410
-
Ime2p and Cdc28p: co-pilots driving meiotic development.J Cell Biochem. 2004 Aug 1;92(5):1025-33. doi: 10.1002/jcb.20131. J Cell Biochem. 2004. PMID: 15258924 Review.
-
Cdc7 protein kinase for DNA metabolism comes of age.Mol Microbiol. 1994 Mar;11(5):805-10. doi: 10.1111/j.1365-2958.1994.tb00358.x. Mol Microbiol. 1994. PMID: 8022258 Review.
Cited by
-
Evolution of Ime2 phosphorylation sites on Cdk1 substrates provides a mechanism to limit the effects of the phosphatase Cdc14 in meiosis.Mol Cell. 2007 Mar 9;25(5):689-702. doi: 10.1016/j.molcel.2007.02.012. Mol Cell. 2007. PMID: 17349956 Free PMC article.
-
Roles of CDK and DDK in Genome Duplication and Maintenance: Meiotic Singularities.Genes (Basel). 2017 Mar 20;8(3):105. doi: 10.3390/genes8030105. Genes (Basel). 2017. PMID: 28335524 Free PMC article. Review.
-
Global alterations of the transcriptional landscape during yeast growth and development in the absence of Ume6-dependent chromatin modification.Mol Genet Genomics. 2015 Oct;290(5):2031-46. doi: 10.1007/s00438-015-1051-5. Epub 2015 May 10. Mol Genet Genomics. 2015. PMID: 25957495 Free PMC article.
-
Dna2 removes toxic ssDNA-RPA filaments generated from meiotic recombination-associated DNA synthesis.Nucleic Acids Res. 2023 Aug 25;51(15):7914-7935. doi: 10.1093/nar/gkad537. Nucleic Acids Res. 2023. PMID: 37351599 Free PMC article.
-
CDC6, a key replication licensing factor, is overexpressed and confers poor prognosis in diffuse large B-cell lymphoma.BMC Cancer. 2023 Oct 13;23(1):978. doi: 10.1186/s12885-023-11186-6. BMC Cancer. 2023. PMID: 37833632 Free PMC article.
References
-
- Aparicio, O.M., Weinstein, D.M., and Bell, S.P. (1997). Components and dynamics of DNA replication complexes in S. cerevisiae: redistribution of MCM proteins and Cdc45p during S phase. Cell 91, 59–69. - PubMed
-
- Calzada, A., Sanchez, M., Sanchez, E., and Bueno, A. (2000). The stability of the Cdc6 protein is regulated by cyclin-dependent kinase/cyclin B complexes in Saccharomyces cerevisiae. J. Biol. Chem. 275, 9734–9741. - PubMed
-
- Cocker, J.H., Piatti, S., Santocanale, C., Nasmyth, K., and Diffley, J.F. (1996). An essential role for the Cdc6 protein in forming the pre-replicative complexes of budding yeast. Nature 379, 180–182. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
