

Phosphorylation of RCC1 in mitosis is essential for producing a high RanGTP concentration on chromosomes and for spindle assembly in mammalian cells
- PMID: 15014043
- PMCID: PMC374234
- DOI: 10.1101/gad.1177304
Phosphorylation of RCC1 in mitosis is essential for producing a high RanGTP concentration on chromosomes and for spindle assembly in mammalian cells
Abstract
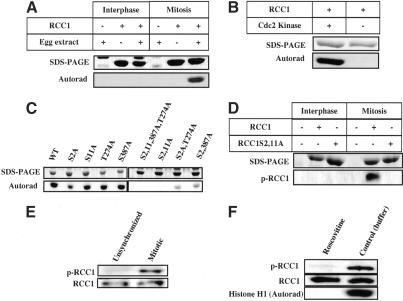
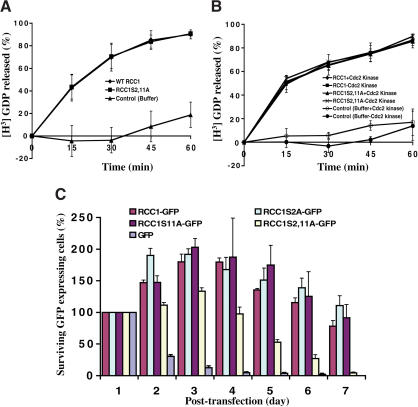
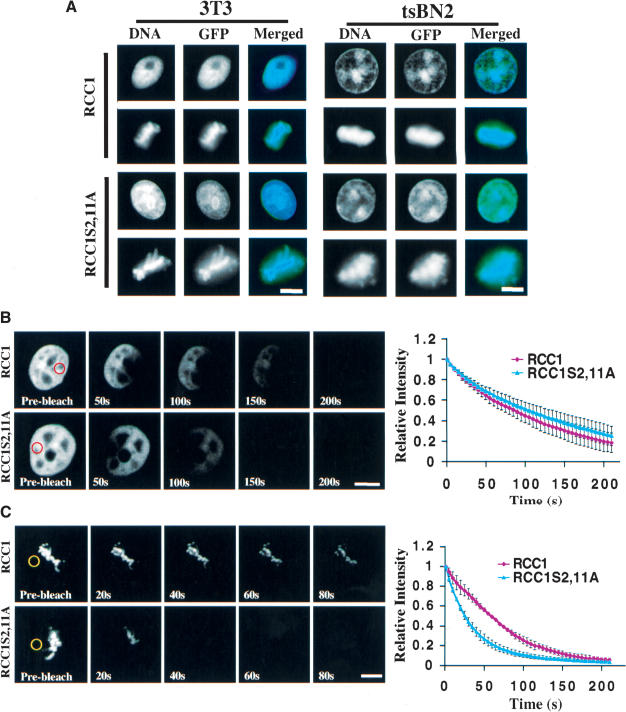
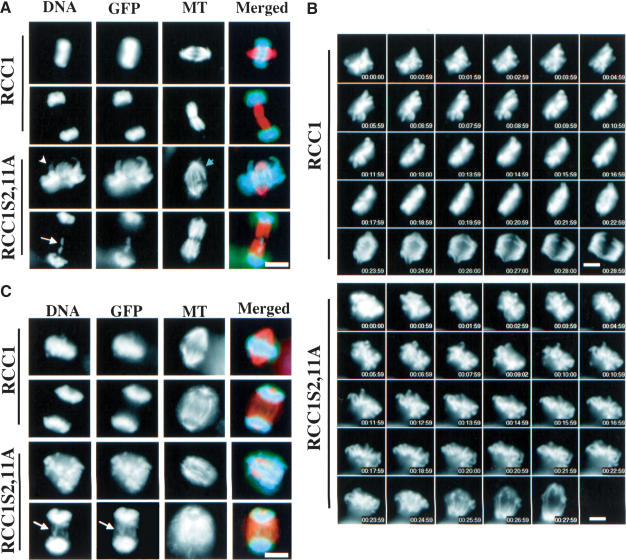
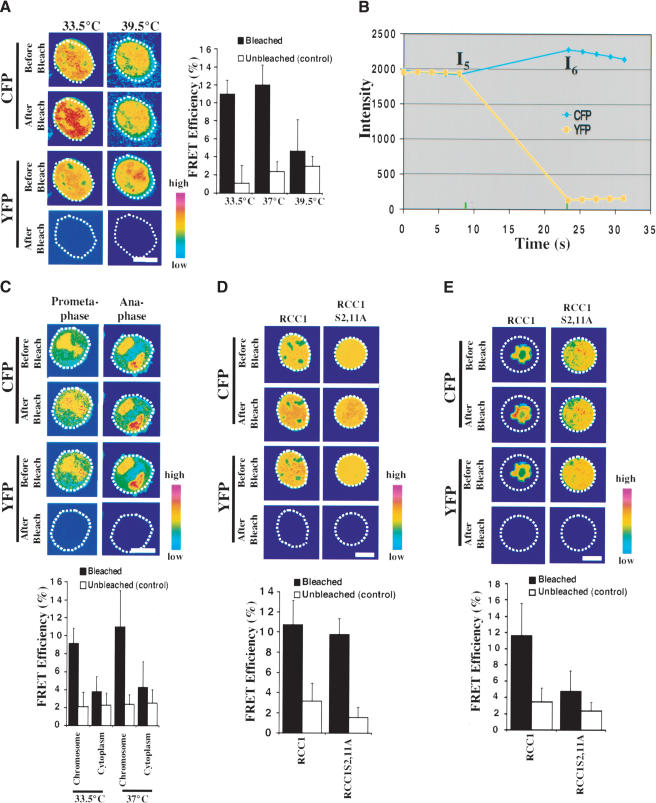
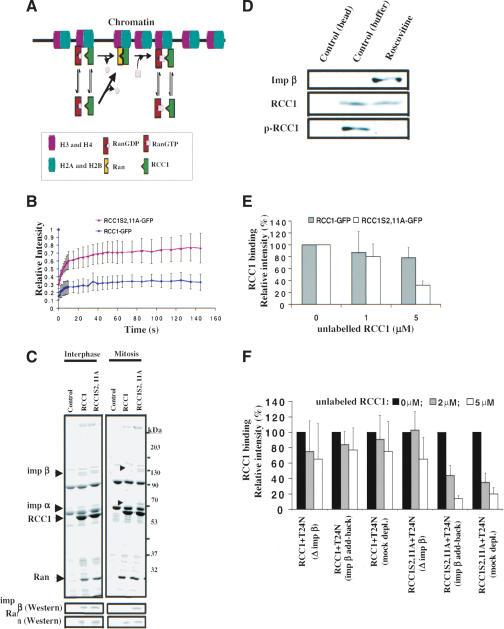
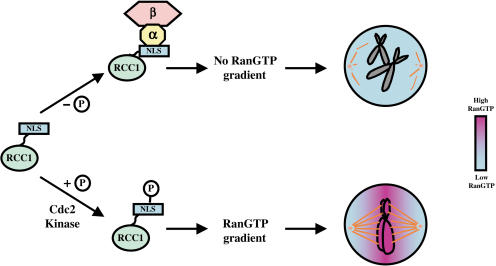
Spindle assembly is subject to the regulatory controls of both the cell-cycle machinery and the Ran-signaling pathway. An important question is how the two regulatory pathways communicate with each other to achieve coordinated regulation in mitosis. We show here that Cdc2 kinase phosphorylates the serines located in or near the nuclear localization signal (NLS) of human RCC1, the nucleotide exchange factor for Ran. This phosphorylation is necessary for RCC1 to generate RanGTP on mitotic chromosomes in mammalian cells, which in turn is required for spindle assembly and chromosome segregation. Moreover, phosphorylation of the NLS of RCC1 is required to prevent the binding of importin alpha and beta to RCC1, thereby allowing RCC1 to couple RanGTP production to chromosome binding. These findings reveal that the cell-cycle machinery directly regulates the Ran-signaling pathway by placing a high RanGTP concentration on the mitotic chromosome in mammalian cells.
Figures
Similar articles
-
Coordination of chromosome alignment and mitotic progression by the chromosome-based Ran signal.Cell Cycle. 2007 Aug 1;6(15):1886-95. doi: 10.4161/cc.6.15.4487. Epub 2007 May 24. Cell Cycle. 2007. PMID: 17671426
-
Phosphorylation regulates the dynamic interaction of RCC1 with chromosomes during mitosis.Curr Biol. 2004 Jun 22;14(12):1099-104. doi: 10.1016/j.cub.2004.05.021. Curr Biol. 2004. PMID: 15203004
-
Targeting of RCC1 to chromosomes is required for proper mitotic spindle assembly in human cells.Curr Biol. 2002 Aug 20;12(16):1442-7. doi: 10.1016/s0960-9822(02)01076-x. Curr Biol. 2002. PMID: 12194828
-
Dynamic changes in nuclear architecture during mitosis: on the role of protein phosphorylation in spindle assembly and chromosome segregation.Exp Cell Res. 1996 Dec 15;229(2):174-80. doi: 10.1006/excr.1996.0356. Exp Cell Res. 1996. PMID: 8986594 Review.
-
The Ran-GTPase and cell-cycle control.Bioessays. 2001 Jan;23(1):77-85. doi: 10.1002/1521-1878(200101)23:1<77::AID-BIES1010>3.0.CO;2-E. Bioessays. 2001. PMID: 11135312 Review.
Cited by
-
Ran is required before metaphase for spindle assembly and chromosome alignment and after metaphase for chromosome segregation and spindle midbody organization.Mol Biol Cell. 2006 Apr;17(4):2069-80. doi: 10.1091/mbc.e05-10-0991. Epub 2006 Feb 15. Mol Biol Cell. 2006. PMID: 16481399 Free PMC article.
-
Phosphorylation by casein kinase 2 regulates Nap1 localization and function.Mol Cell Biol. 2008 Feb;28(4):1313-25. doi: 10.1128/MCB.01035-07. Epub 2007 Dec 17. Mol Cell Biol. 2008. PMID: 18086883 Free PMC article.
-
Ran GTPase guanine nucleotide exchange factor RCC1 is phosphorylated on serine 11 by cdc2 kinase in vitro.Mol Biol Rep. 2009 Apr;36(4):717-23. doi: 10.1007/s11033-008-9234-3. Epub 2008 Jun 21. Mol Biol Rep. 2009. PMID: 18568422
-
Nuclear reformation after mitosis requires downregulation of the Ran GTPase effector RanBP1 in mammalian cells.Chromosoma. 2010 Dec;119(6):651-68. doi: 10.1007/s00412-010-0286-5. Epub 2010 Jul 24. Chromosoma. 2010. PMID: 20658144
-
Remodeling of the pioneer translation initiation complex involves translation and the karyopherin importin beta.Genes Dev. 2009 Nov 1;23(21):2537-50. doi: 10.1101/gad.1817109. Genes Dev. 2009. PMID: 19884259 Free PMC article.
References
-
- Antonio C., Ferby, I., Wilhelm, H., Jones, M., Karsenti, E., Nebreda, A.R., and Vernos, I. 2000. Xkid, a chromokinesin required for chromosome alignment on the metaphase plate. Cell 102: 425–435. - PubMed
-
- Arnaoutov A. and Dasso, M. 2003. The Ran GTPase regulates kinetochore function. Dev Cell. 5: 99–111. - PubMed
-
- Bilbao-Cortes D., Hetzer, M., Langst, G., Becker, P.B., and Mattij, I.W. 2002. Ran binds to chromatin by two distinct mechanisms. Curr. Biol. 12: 1151–1156. - PubMed
-
- Carazo-Salas R.E., Guarguaglini, G., Gruss, O.J., Segref, A., Karsenti, E., and Mattaj, I.W. 1999. Generation of GTP-bound Ran by RCC1 is required for chromatin-induced mitotic spindle formation. Nature 400: 178–181. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous