

Cellular targets and mechanisms of nitros(yl)ation: an insight into their nature and kinetics in vivo
- PMID: 15014175
- PMCID: PMC384737
- DOI: 10.1073/pnas.0306706101
Cellular targets and mechanisms of nitros(yl)ation: an insight into their nature and kinetics in vivo
Abstract
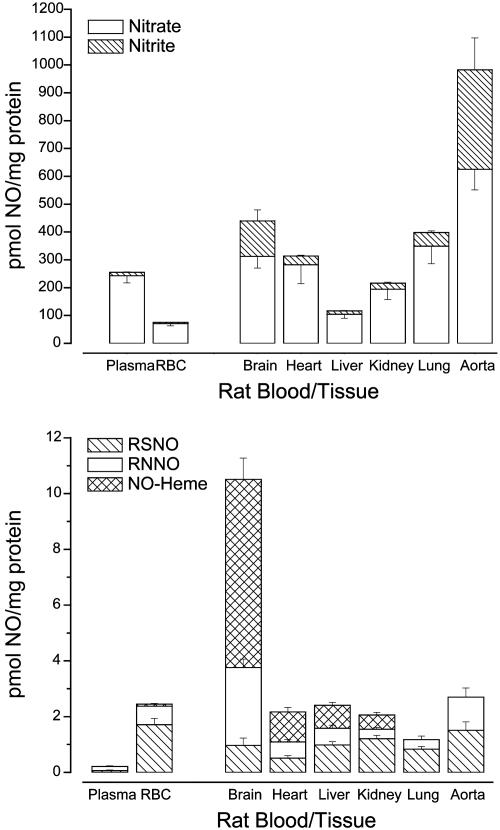
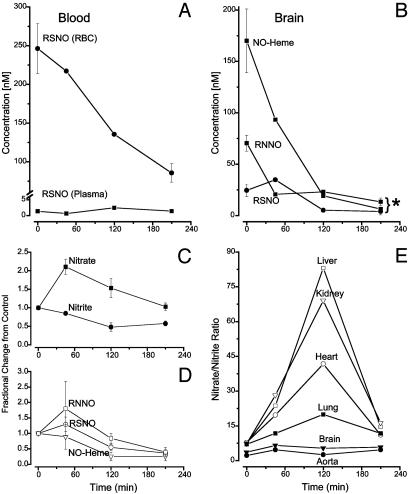
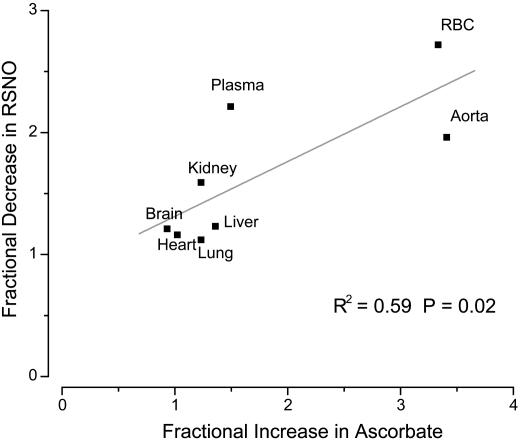
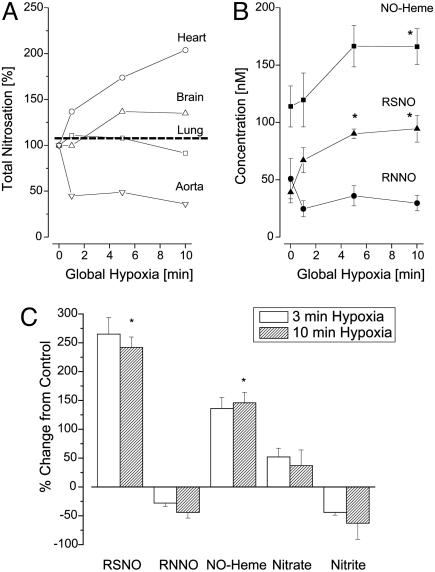
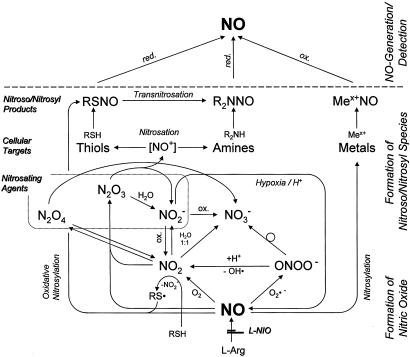
There is mounting evidence that the established paradigm of nitric oxide (NO) biochemistry, from formation through NO synthases, over interaction with soluble guanylyl cyclase, to eventual disposal as nitrite/nitrate, represents only part of a richer chemistry through which NO elicits biological signaling. Additional pathways have been suggested that include interaction of NO-derived metabolites with thiols and metals to form S-nitrosothiols (RSNOs) and metal nitrosyls. Despite the overwhelming attention paid in this regard to RSNOs, little is known about the stability of these species, their significance outside the circulation, and whether other nitros(yl)ation products are of equal importance. We here show that N-nitrosation and heme-nitrosylation are indeed as ubiquitous as S-nitrosation in vivo and that the products of these reactions are constitutively present throughout the organ system. Our study further reveals that all NO-derived products are highly dynamic, have fairly short lifetimes, and are linked to tissue oxygenation and redox state. Experimental evidence further suggests that nitroso formation occurs substantially by means of oxidative nitrosylation rather than NO autoxidation, explaining why S-nitrosation can compete effectively with nitrosylation. Moreover, tissue nitrite can serve as a significant extravascular pool of NO during brief periods of hypoxia, and tissue nitrate/nitrite ratios can serve as indicators of the balance between local oxidative and nitrosative stress. These findings vastly expand our understanding of the fate of NO in vivo and provide a framework for further exploration of the significance of nitrosative events in redox sensing and signaling. The findings also raise the intriguing possibility that N-nitrosation is directly involved in the modulation of protein function.
Figures
References
-
- Moncada, S., Palmer, R. M. J. &. Higgs, E. A. (1991) Pharmacol. Rev. 43, 109-142. - PubMed
-
- Stamler, J. S., Singel, D. J. & Loscalzo, J. (1992) Science 258, 1898-1902. - PubMed
-
- Foster, M. W., McMahon, T. J. & Stamler, J. S. (2003) Trends Mol. Med. 9, 160-168. - PubMed
-
- Espey, M. G., Miranda, K. M., Thomas, D. D. & Wink, D. A. (2001) J. Biol. Chem. 276, 30085-30091. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
