

Mechanism controlling perpendicular alignment of the spindle to the axis of cell division in fission yeast
- PMID: 15014440
- PMCID: PMC381420
- DOI: 10.1038/sj.emboj.7600156
Mechanism controlling perpendicular alignment of the spindle to the axis of cell division in fission yeast
Abstract
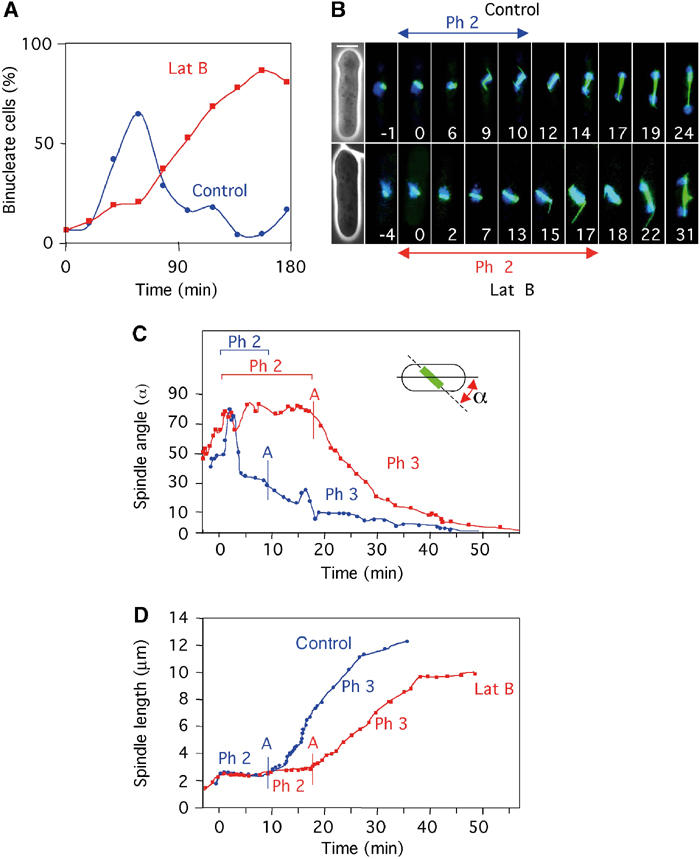
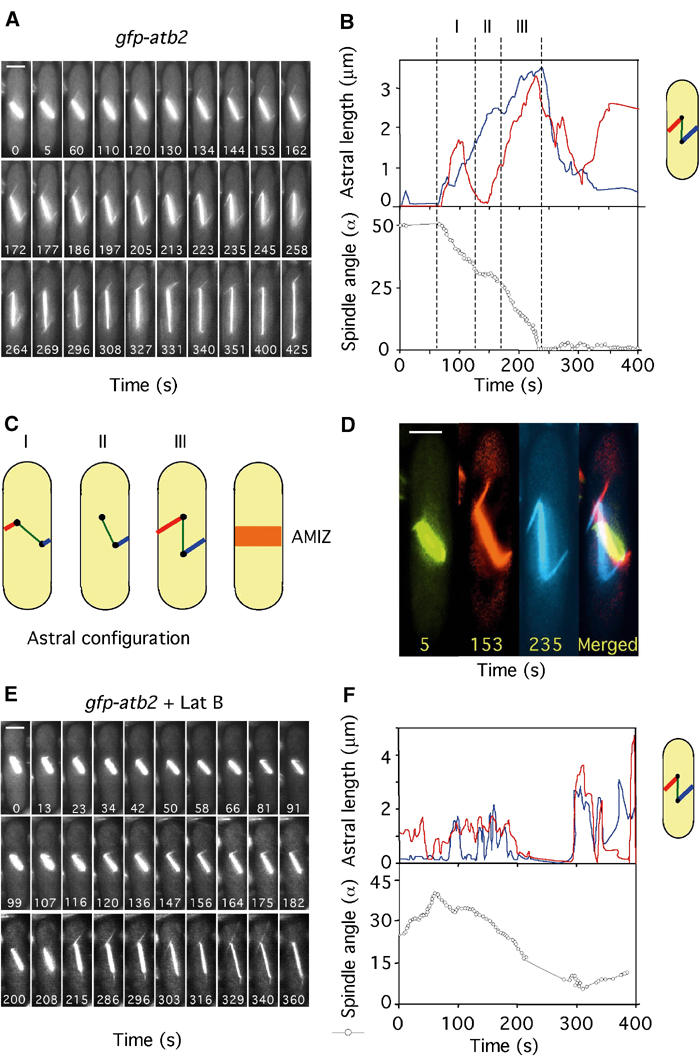
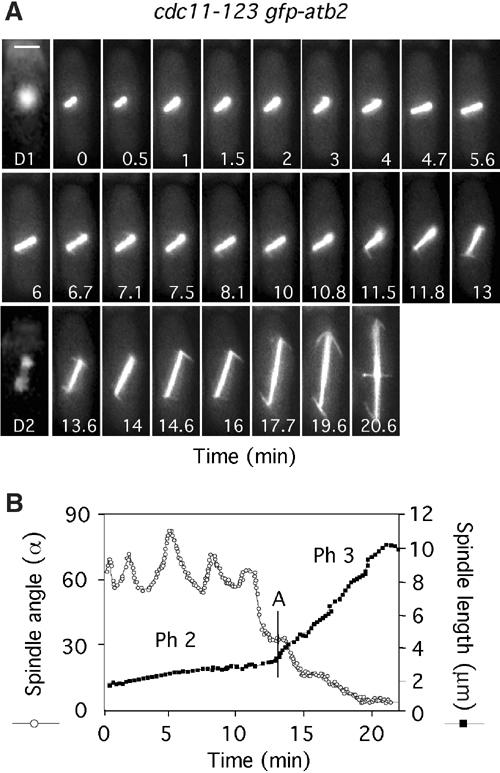
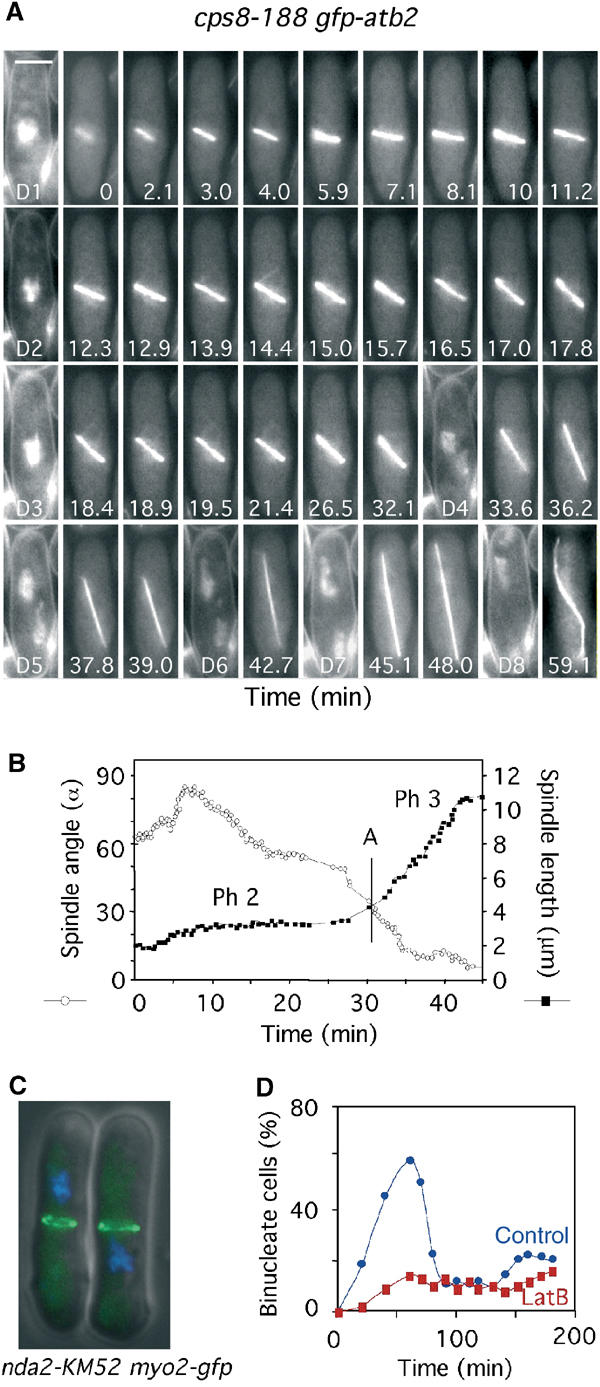
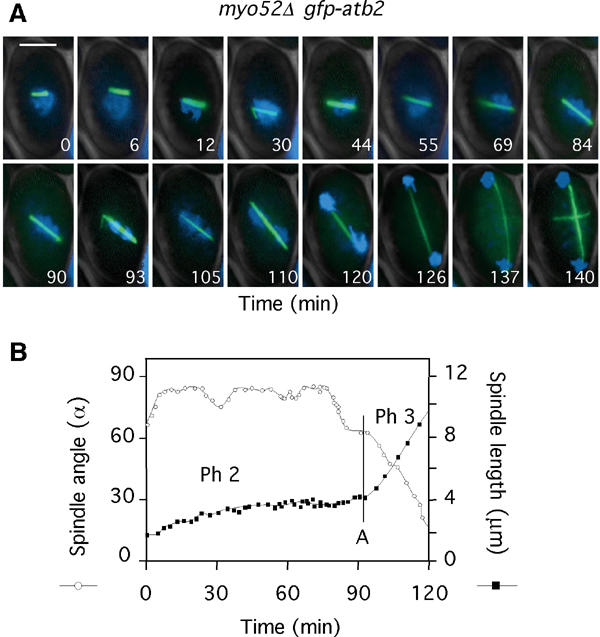
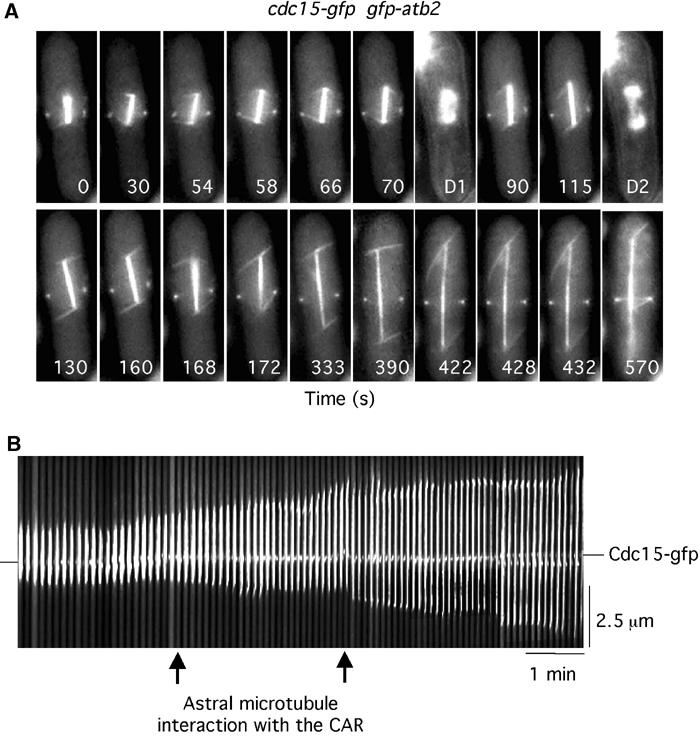
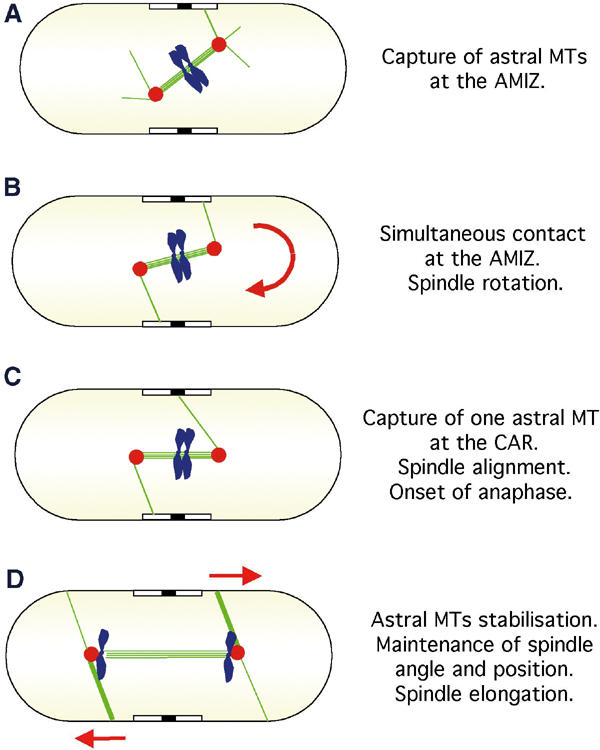
In animal cells, the mitotic spindle is aligned perpendicular to the axis of cell division. This ensures that sister chromatids are separated to opposite sides of the cytokinetic actomyosin ring (CAR). We show that, in fission yeast, spindle rotation is dependent on the interaction of astral microtubules with the cortical actin cytoskeleton. Interaction initially occurs with a region surrounding the nucleus, which we term the astral microtubule interaction zone (AMIZ). Simultaneous contact of astral microtubules from both poles with the AMIZ directs spindle rotation and this requires both actin and two type V myosins, Myo51 and Myo52. Astral microtubules from one pole only then contact the CAR, which is located at the centre of the AMIZ. We demonstrate that the anillin homologue Mid1, which dictates correct placement of the CAR, is necessary to stabilise the mitotic spindle perpendicular to the axis of cell division. Finally, we show that the position of the mitotic spindle is monitored by a checkpoint that regulates the timing of sister chromatid separation.
Figures

References
-
- Aist JR, Bayles CJ, Tao W, Berns MW (1991) Direct experimental evidence for the existence, structural basis and function of astral forces during anaphase B in vivo. J Cell Sci 100: 279–288 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
