

Haplotype diversity across 100 candidate genes for inflammation, lipid metabolism, and blood pressure regulation in two populations
- PMID: 15015130
- PMCID: PMC1181939
- DOI: 10.1086/382227
Haplotype diversity across 100 candidate genes for inflammation, lipid metabolism, and blood pressure regulation in two populations
Abstract
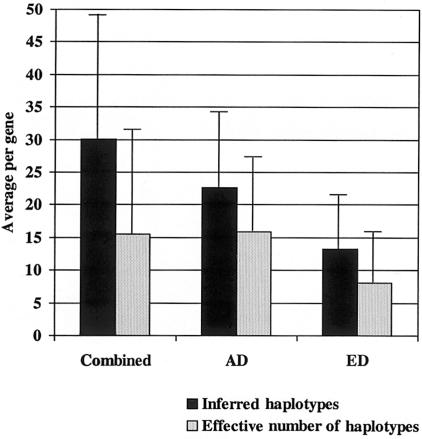
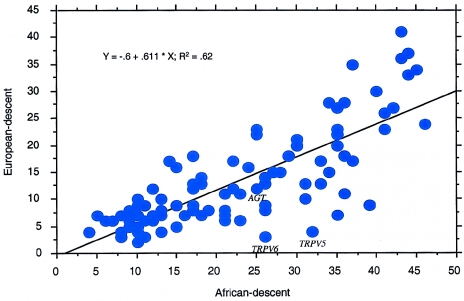
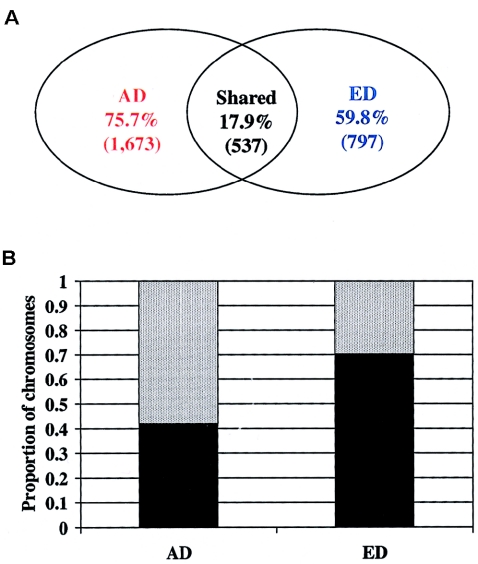
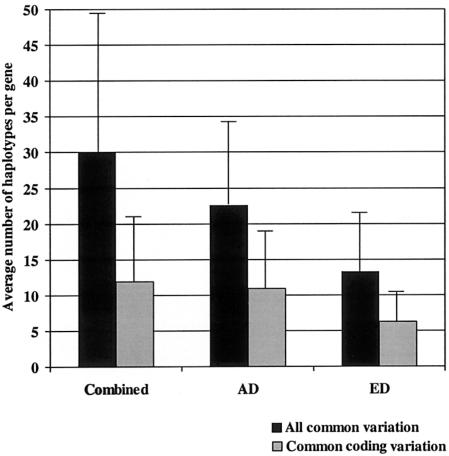
Recent studies have suggested that a significant fraction of the human genome is contained in blocks of strong linkage disequilibrium, ranging from ~5 to >100 kb in length, and that within these blocks a few common haplotypes may account for >90% of the observed haplotypes. Furthermore, previous studies have suggested that common haplotypes in candidate genes are generally shared across populations and represent the majority of chromosomes in each population. The conclusions drawn from these preliminary studies, however, are based on an incomplete knowledge of the variation in the regions examined. To bridge this gap in knowledge, we have completely resequenced 100 candidate genes in a population of African descent and one of European descent. Although these genes have been well studied because of their medical importance, we demonstrate that a large amount of sequence variation has not yet been described. We also report that the average number of inferred haplotypes per gene, when complete data is used, is higher than in previous reports and that the number and proportion of all haplotypes represented by common haplotypes per gene is variable. Furthermore, we demonstrate that haplotypes shared between the two populations constitute only a fraction of the total number of haplotypes observed and that these shared haplotypes represent fewer of the African-descent chromosomes than was expected from previous studies. Finally, we show that restricting variation discovery to coding regions does not adequately describe all common haplotypes or the true haplotype block structure observed when all common variation is used to infer haplotypes. These data, derived from complete knowledge of genetic variation in these genes, suggest that the haplotype architecture of candidate genes across the human genome is more complex than previously suggested, with important implications for candidate gene and genomewide association studies.
Figures
References
Electronic-Database Information
-
- GenBank, http://www.ncbi.nlm.nih.gov/Genbank/ (accession numbers for all genes are listed in [online only])
-
- HaploBlockFinder V0.6b, http://cgi.uc.edu/cgi-bin/kzhang/haploBlockFinder.cgi/
-
- Nickerson Group, http://droog.gs.washington.edu
-
- Pharmacogenetics Research Network, http://www.nigms.nih.gov/pharmacogenetics/
References
-
- Cargill M, Altshuler D, Ireland J, Sklar P, Ardlie K, Patil N, Shaw N, Lane CR, Lim EP, Kalyanaraman N, Nemesh J, Ziaugra L, Friedland L, Rolfe A, Warrington J, Lipshutz R, Daley GQ, Lander ES (1999) Characterization of single-nucleotide polymorphisms in coding regions of human genes. Nat Genet 22:231–23810.1038/10290 - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
