

Review
doi: 10.1016/j.virol.2003.11.029.
The relevance of complement to virus biology
Affiliations
- PMID: 15015499
- PMCID: PMC7172316
- DOI: 10.1016/j.virol.2003.11.029
Item in Clipboard
Review
The relevance of complement to virus biology
Virology.
.
No abstract available
Figures
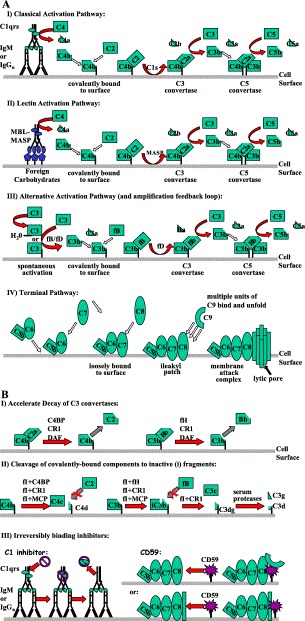
Activation pathways of complement (A) and their regulation (B). (A) Three separate routes of complement activation are depicted (I–III). Common to all three pathways is the formation of C3 and C5 convertase enzyme complexes. The C3 convertase (C4b2a or C3bBb) cleaves C3 into C3a (chemotaxin) and C3b. The latter protein forms the C5 convertase, which cleaves C5 to C5a (anaphylatoxin) and C5b. The production of C5b enables the formation of a lytic pore, known as the membrane attack complex (MAC). (I) Activation of the classical pathway results from the binding of C1q to two immunoglobulin Fc regions of antigen-bound IgG and IgM. This binding induces the autolytic cleavage of C1r and the subsequent cleavage of C1s to its active from. Activated C1s cleaves C4. The resulting C4b then binds covalently to the cell surface enabling C2 to bind. C1s then cleaves C2 complexed with C4b, generating the classical C3 convertase, C4b2a. (II) The lectin pathway is triggered in an identical manner except that it is recognized by the mannan-binding lectin (MBL) of foreign carbohydrate that activates the MBL-associated serine proteases (MASP), which are capable of cleaving C4 and C2 to create the C3 convertase, C4b2a. While three separate MASPs have been identified, only MASP-2 can form a C3 convertase (reviewed in Schwaeble et al., 2002). (III) The alternative pathway is continually active, but only amplifies on an activating surface because of insufficient regulation. It also serves as a C3 convertase amplification loop for the other two pathways. The alternative pathway is triggered by the constant low-level spontaneous cleavage of C3 that occurs by nonspecific release of the internal thioester bond. The resulting hydrolysed C3 (either fluid phase or cell-bound) then forms a complex with factor B (fB), which is itself cleaved by serum protease factor D (fD). This complex then generates more C3b and results in the formation of the alternative pathway C3 convertase (C3bBb), which is further stabilised by the association of the serum protein properdin (not shown). (IV) C5b, generated by any of the previous activation pathways, associates noncovalently with C6. This association enables a loose interaction with the membrane surface, which is strengthened by the subsequent noncovalent association of C7 and C8 that causes insertion of the complex into the membrane. The full membrane attack complex consists of a lytic pore formed by the further incorporation of 12–16 molecules of C9. (B) Complement is regulated by several mechanisms (see also Table 1): (I) surface-bound and serum proteins accelerate the decay of the C3 convertases, in many cases, (II) subsequently inducing factor I (fI) cleavage of the covalently attached components of the C3 convertase to fragments that can no longer bind C2 (see AI and AII) or fB (see AIII). (III) Conversely, other regulators act through “suicide” irreversible association with either the terminal complement components (CD59) or the initial C1qrs complex of the classical pathway (C1 inhibitor) resulting in subsequent removal of the C1rs protease complex.
References
-
- Albrecht J.C., Nicholas J., Cameron K.R., Newman C., Fleckenstein B., Honess R.W. Herpesvirus saimiri has a gene specifying a homologue of the cellular membrane glycoprotein CD59. Virology. 1992;190(1):527–530. - PubMed
-
- Antonsson A., Johansson P.J. Binding of human and animal immunoglobulins to the IgG Fc receptor induced by human cytomegalovirus. J. Gen. Virol. 2001;82(Pt 5):1137–1145. - PubMed
-
- Carney D.F., Koski C.L., Shin M.L. Elimination of terminal complement intermediates from the plasma membrane of nucleated cells: the rate of disappearance differs for cells carrying C5b-7 or C5b-8 or a mixture of C5b-8 with a limited number of C5b-9. J. Immunol. 1985;134(3):1804–1809. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
