

Azido-containing diketo acid derivatives inhibit human immunodeficiency virus type 1 integrase in vivo and influence the frequency of deletions at two-long-terminal-repeat-circle junctions
- PMID: 15016842
- PMCID: PMC371038
- DOI: 10.1128/jvi.78.7.3210-3222.2004
Azido-containing diketo acid derivatives inhibit human immunodeficiency virus type 1 integrase in vivo and influence the frequency of deletions at two-long-terminal-repeat-circle junctions
Abstract
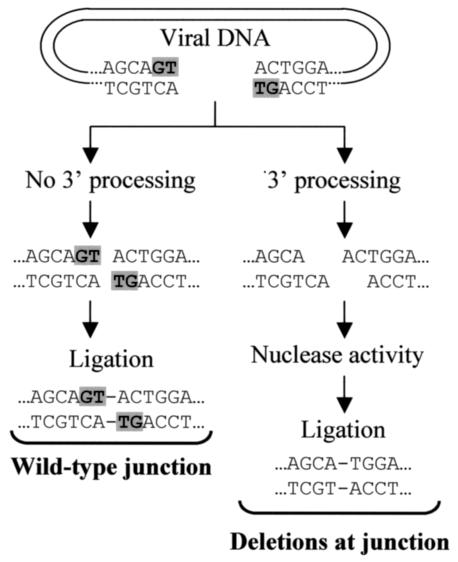
We previously found that azido-containing beta-diketo acid derivatives (DKAs) are potent inhibitors of human immunodeficiency virus type 1 (HIV-1) integrase (IN) (X. Zhang et al., Bioorg. Med. Chem. Lett., 13:1215-1219, 2003). To characterize the intracellular mechanisms of action of DKAs, we analyzed the antiviral activities of two potent azido-containing DKAs with either a monosubstitution or a disubstitution of azido groups, using single- and multiple-replication-cycle assays. Both azido-containing DKAs significantly inhibited HIV-1 infection in 293T, CEM-SS, and H9 cells (50% inhibitory concentration = 2 to 13 micro M) and exhibited low cytotoxicity (50% cytotoxic concentration = 60 to 600 micro M). Inhibition of HIV-1 IN in vivo was demonstrated by the observation that previously described L-708,906 resistance mutations in HIV-1 IN (T66I and T66I/S153Y) also conferred resistance to the azido-group-containing DKAs. In vitro assays and in vivo analysis indicated that the DKAs did not significantly inhibit the 3' processing and selectively inhibited the strand transfer reaction. In addition, quantitative PCR indicated that two-long-terminal-repeat (2-LTR) circles were elevated in the presence of the azido-containing DKAs, confirming that HIV-1 IN was the intracellular target of viral inhibition. To gain insight into the mechanism by which the DKAs increased 2-LTR-circle formation of 3'-processed viral DNAs, we performed extensive DNA sequencing analysis of 2-LTR-circle junctions. The results indicated that the frequency of deletions at the circle junctions was elevated from 19% for the untreated controls to 32 to 41% in the presence of monosubstituted (but not disubstituted) DKAs. These results indicate that the structure of the DKAs can influence the extent of degradation of viral DNA ends by host nucleases and the frequency of deletions at the 2-LTR-circle junctions. Thus, sequencing analysis of 2-LTR-circle junctions can elucidate the intracellular mechanisms of action of HIV-1 IN inhibitors.
Figures






Similar articles
-
Inhibitors of strand transfer that prevent integration and inhibit HIV-1 replication in cells.Science. 2000 Jan 28;287(5453):646-50. doi: 10.1126/science.287.5453.646. Science. 2000. PMID: 10649997
-
Human immunodeficiency virus type 1 (HIV-1) integrase: resistance to diketo acid integrase inhibitors impairs HIV-1 replication and integration and confers cross-resistance to L-chicoric acid.J Virol. 2004 Jun;78(11):5835-47. doi: 10.1128/JVI.78.11.5835-5847.2004. J Virol. 2004. PMID: 15140981 Free PMC article.
-
Dihydroxythiophenes are novel potent inhibitors of human immunodeficiency virus integrase with a diketo acid-like pharmacophore.J Virol. 2006 Jul;80(14):6883-94. doi: 10.1128/JVI.00306-06. J Virol. 2006. PMID: 16809294 Free PMC article.
-
Characterization and structural analysis of HIV-1 integrase conservation.AIDS Rev. 2009 Jan-Mar;11(1):17-29. AIDS Rev. 2009. PMID: 19290031 Review.
-
Resistance to HIV-1 integrase inhibitors: A structural perspective.Drug Resist Updat. 2010 Aug-Oct;13(4-5):139-50. doi: 10.1016/j.drup.2010.05.001. Epub 2010 Jun 8. Drug Resist Updat. 2010. PMID: 20570551 Review.
Cited by
-
Efficient and specific internal cleavage of a retroviral palindromic DNA sequence by tetrameric HIV-1 integrase.PLoS One. 2007 Jul 11;2(7):e608. doi: 10.1371/journal.pone.0000608. PLoS One. 2007. PMID: 17622353 Free PMC article.
-
Integrase and integration: biochemical activities of HIV-1 integrase.Retrovirology. 2008 Dec 17;5:114. doi: 10.1186/1742-4690-5-114. Retrovirology. 2008. PMID: 19091057 Free PMC article. Review.
-
A large U3 deletion causes increased in vivo expression from a nonintegrating lentiviral vector.Mol Ther. 2008 Dec;16(12):1968-76. doi: 10.1038/mt.2008.199. Epub 2008 Sep 16. Mol Ther. 2008. PMID: 18797449 Free PMC article.
-
A targeted DNA substrate mechanism for the inhibition of HIV-1 integrase by inhibitors with antiretroviral activity.FEBS Open Bio. 2016 Feb 24;6(4):234-50. doi: 10.1002/2211-5463.12025. eCollection 2016 Apr. FEBS Open Bio. 2016. PMID: 27239438 Free PMC article.
-
Dominant Negative MA-CA Fusion Protein Is Incorporated into HIV-1 Cores and Inhibits Nuclear Entry of Viral Preintegration Complexes.J Virol. 2019 Oct 15;93(21):e01118-19. doi: 10.1128/JVI.01118-19. Print 2019 Nov 1. J Virol. 2019. PMID: 31413124 Free PMC article.
References
-
- Beale, K. K., and W. E. Robinson, Jr. 2000. Combinations of reverse transcriptase, protease, and integrase inhibitors can be synergistic in vitro against drug-sensitive and RT inhibitor-resistant molecular clones of HIV-1. Antivir. Res. 46:223-232. - PubMed
-
- Billich, A. 2003. S-1360 Shionogi-GlaxoSmithKline. Curr. Opin. Investig. Drugs 4:206-209. - PubMed
-
- Brin, E., J. Yi, A. M. Skalka, and J. Leis. 2000. Modeling the late steps in HIV-1 retroviral integrase-catalyzed DNA integration. J. Biol. Chem. 275:39287-39295. - PubMed
-
- Burke, T. R., Jr., M. R. Fesen, A. Mazumder, J. Wang, A. M. Carothers, D. Grunberger, J. Driscoll, K. Kohn, and Y. Pommier. 1995. Hydroxylated aromatic inhibitors of HIV-1 integrase. J. Med. Chem. 38:4171-4178. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
