

PU.1 binding to ets motifs within the equine infectious anemia virus long terminal repeat (LTR) enhancer: regulation of LTR activity and virus replication in macrophages
- PMID: 15016863
- PMCID: PMC371083
- DOI: 10.1128/jvi.78.7.3407-3418.2004
PU.1 binding to ets motifs within the equine infectious anemia virus long terminal repeat (LTR) enhancer: regulation of LTR activity and virus replication in macrophages
Abstract
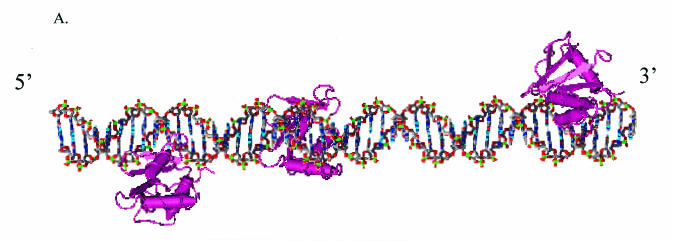
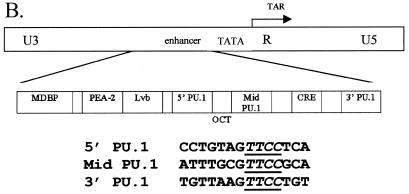
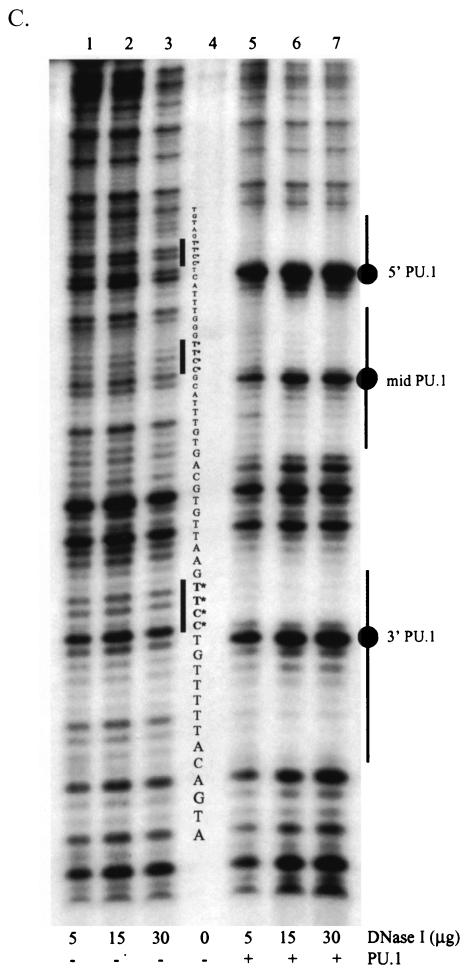
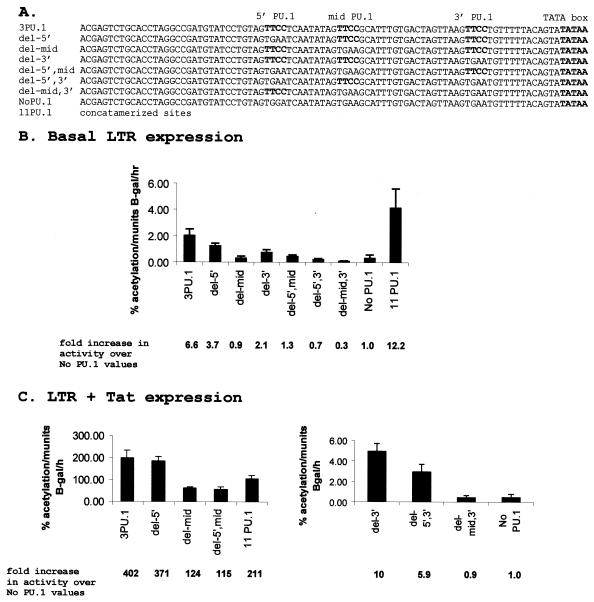
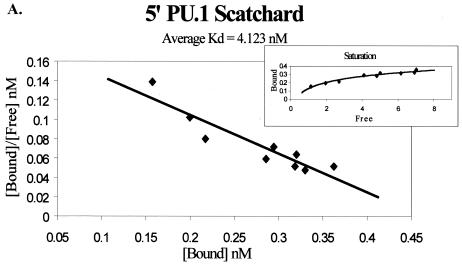
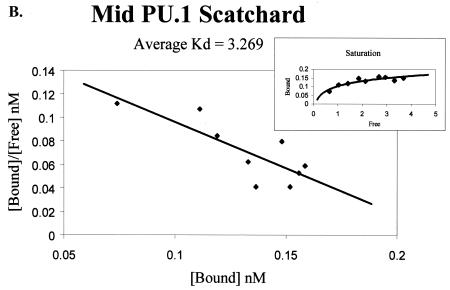
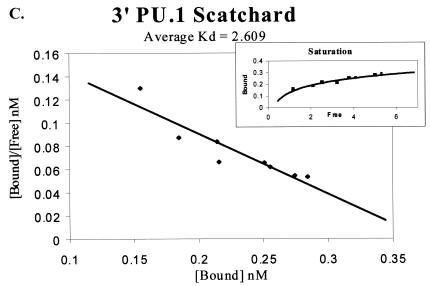
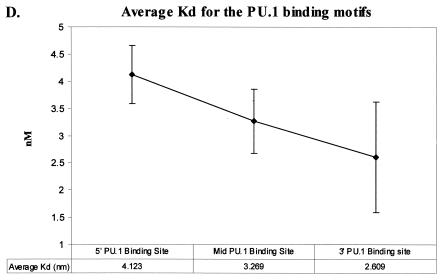
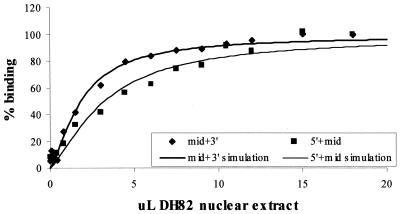
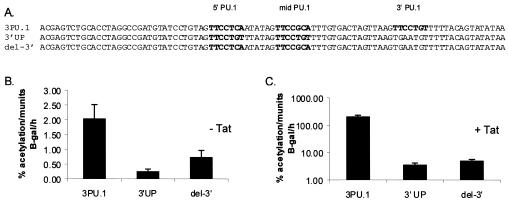
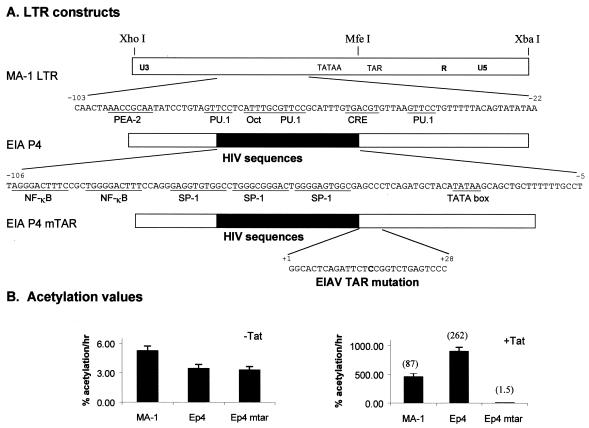
Binding of the transcription factor PU.1 to its DNA binding motif regulates the expression of a number of B-cell- and myeloid-specific genes. The long terminal repeat (LTR) of macrophage-tropic strains of equine infectious anemia virus (EIAV) contains three PU.1 binding sites, namely an invariant promoter-proximal site as well as two upstream sites. We have previously shown that these sites are important for EIAV LTR activity in primary macrophages (W. Maury, J. Virol. 68:6270-6279, 1994). Since the sequences present in these three binding motifs are not identical, we sought to determine the role of these three sites in EIAV LTR activity. While DNase I footprinting studies indicated that all three sites within the enhancer were bound by recombinant PU.1, reporter gene assays demonstrated that the middle motif was most important for basal levels of LTR activity in macrophages and that the 5' motif had little impact. The impact of the 3' site became evident in Tat transactivation studies, in which the loss of the site reduced Tat-transactivated expression 40-fold. In contrast, elimination of the 5' site had no effect on Tat-mediated activity. Binding studies were performed to determine whether differences in PU.1 binding affinity for the three sites correlated with the relative impact of each site on LTR transcription. While small differences were observed in the binding affinities of the three sites, with the promoter-proximal site having the strongest binding affinity, these differences could not account for the dramatic differences observed in the transcriptional effects. Instead, the promoter-proximal position of the 3' motif appeared to be critical for its transcriptional impact and suggested that the PU.1 sites may serve different roles depending upon the location of the sites within the enhancer. Infectivity studies demonstrated that an LTR containing an enhancer composed of the three PU.1 sites was not sufficient to drive viral replication in macrophages. These findings indicate that while the promoter-proximal PU.1 site is the most critical site for EIAV LTR activity in the presence of Tat, other elements within the enhancer are needed for EIAV replication in macrophages.
Figures
Similar articles
-
Effects of long terminal repeat sequence variation on equine infectious anemia virus replication in vitro and in vivo.Virology. 1999 Oct 25;263(2):408-17. doi: 10.1006/viro.1999.9921. Virology. 1999. PMID: 10544113
-
Cell specificity of the transcription-factor repertoire used by a lentivirus: motifs important for expression of equine infectious anemia virus in nonmonocytic cells.Virology. 2000 Feb 15;267(2):267-78. doi: 10.1006/viro.1999.0144. Virology. 2000. PMID: 10662622
-
Protein interactions with DNA elements in variant equine infectious anemia virus enhancers and their impact on transcriptional activity.J Virol. 1993 Nov;67(11):6586-95. doi: 10.1128/JVI.67.11.6586-6595.1993. J Virol. 1993. PMID: 8411361 Free PMC article.
-
The PU.1/Spi-1 proto-oncogene is a transcriptional regulator of a lentivirus promoter.J Virol. 1993 Jul;67(7):3885-90. doi: 10.1128/JVI.67.7.3885-3890.1993. J Virol. 1993. PMID: 8389910 Free PMC article.
-
Cellular specificity of HIV-1 replication can be controlled by LTR sequences.Virology. 2003 Sep 30;314(2):680-95. doi: 10.1016/s0042-6822(03)00508-7. Virology. 2003. PMID: 14554095
Cited by
-
Transcription factor binding sites in the pol gene intragenic regulatory region of HIV-1 are important for virus infectivity.Nucleic Acids Res. 2005 Aug 1;33(13):4285-310. doi: 10.1093/nar/gki720. Print 2005. Nucleic Acids Res. 2005. PMID: 16061936 Free PMC article.
-
Endocytosis and a low-pH step are required for productive entry of equine infectious anemia virus.J Virol. 2005 Dec;79(23):14482-8. doi: 10.1128/JVI.79.23.14482-14488.2005. J Virol. 2005. PMID: 16282447 Free PMC article.
-
Effects of random mutations in the human immunodeficiency virus type 1 transcriptional promoter on viral fitness in different host cell environments.J Virol. 2006 Jul;80(13):6678-85. doi: 10.1128/JVI.02547-05. J Virol. 2006. PMID: 16775355 Free PMC article.
-
Evolution of the equine infectious anemia virus long terminal repeat during the alteration of cell tropism.J Virol. 2005 May;79(9):5653-64. doi: 10.1128/JVI.79.9.5653-5664.2005. J Virol. 2005. PMID: 15827180 Free PMC article.
-
Genetic variation in the long terminal repeat associated with the transition of Chinese equine infectious anemia virus from virulence to avirulence.Virus Genes. 2009 Apr;38(2):285-8. doi: 10.1007/s11262-008-0317-y. Epub 2009 Jan 7. Virus Genes. 2009. PMID: 19130201
References
-
- Barboric, M., R. M. Nissen, S. Kanazawa, N. Jabrane-Ferrat, and B. M. Peterlin. 2001. NF-kappaB binds P-TEFb to stimulate transcriptional elongation by RNA polymerase II. Mol. Cell 8:327-337. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
