

Sequence-specific inhibition of small RNA function
- PMID: 15024405
- PMCID: PMC350664
- DOI: 10.1371/journal.pbio.0020098
Sequence-specific inhibition of small RNA function
Abstract
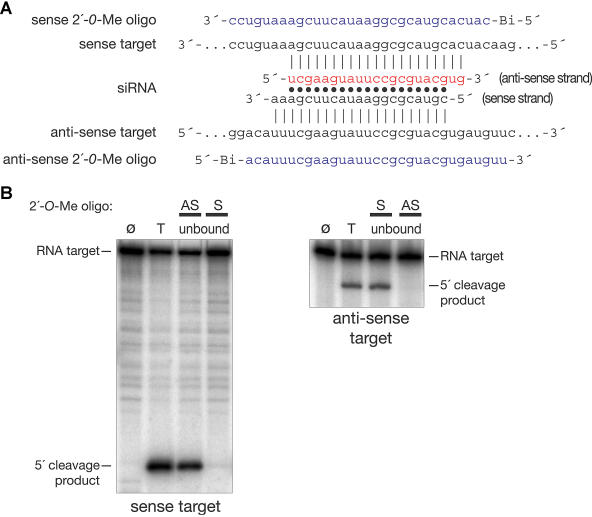
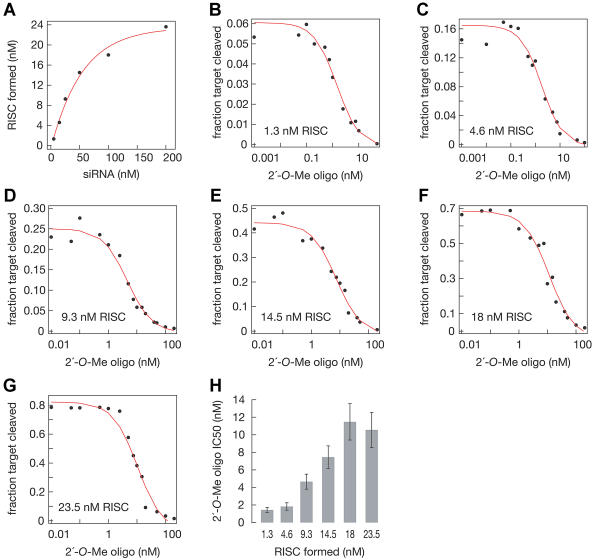
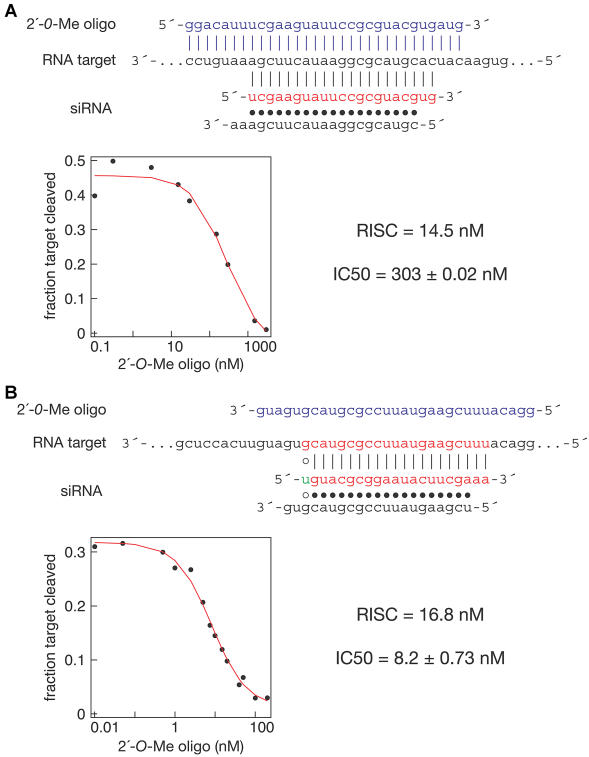
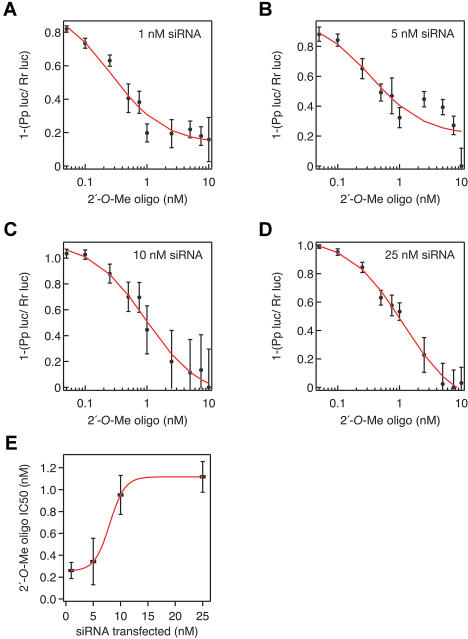
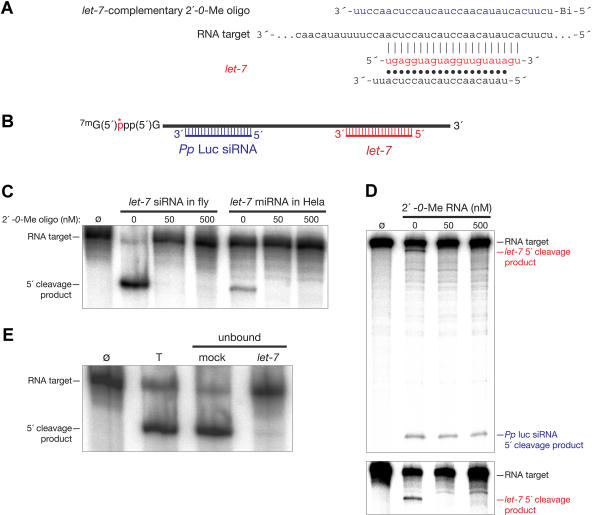
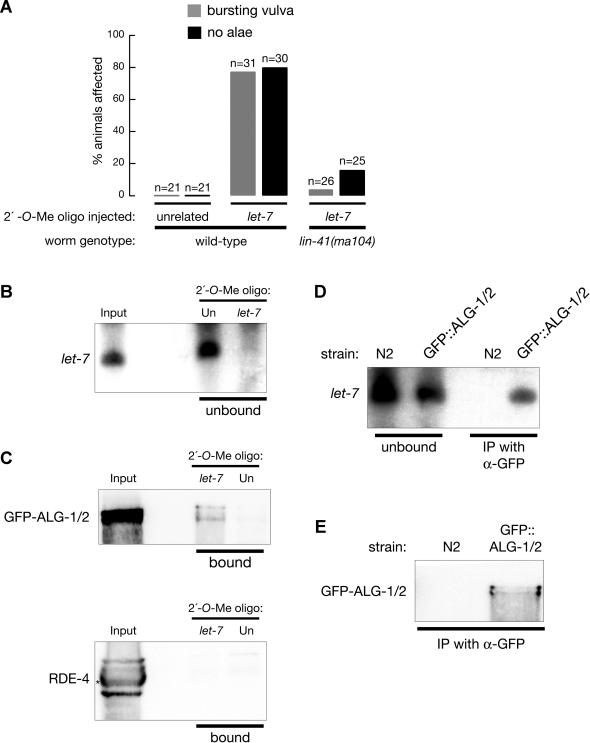
Hundreds of microRNAs (miRNAs) and endogenous small interfering RNAs (siRNAs) have been identified from both plants and animals, yet little is known about their biochemical modes of action or biological functions. Here we report that 2'-O-methyl oligonucleotides can act as irreversible, stoichiometric inhibitors of small RNA function. We show that a 2'-O-methyl oligonucleotide complementary to an siRNA can block mRNA cleavage in Drosophila embryo lysates and HeLa cell S100 extracts and in cultured human HeLa cells. In Caenorhabditis elegans, injection of the 2'-O-methyl oligonucleotide complementary to the miRNA let-7 can induce a let-7 loss-of-function phenocopy. Using an immobilized 2'-O-methyl oligonucleotide, we show that the C. elegans Argonaute proteins ALG-1 and ALG-2, which were previously implicated in let-7 function through genetic studies, are constituents of a let-7-containing protein-RNA complex. Thus, we demonstrate that 2'-O-methyl RNA oligonucleotides can provide an efficient and straightforward way to block small RNA function in vivo and furthermore can be used to identify small RNA-associated proteins that mediate RNA silencing pathways.
Conflict of interest statement
The authors have declared that no conflicts of interest exist.
Figures
References
-
- Ambros V, Lee RC, Lavanway A, Williams PT, Jewell D. MicroRNAs and other tiny endogenous RNAs in C. elegans . Curr Biol. 2003;13:807–818. - PubMed
-
- Aravin AA, Lagos-Quintana M, Yalcin A, Zavolan M, Marks D, et al. The small RNA profile during Drosophila melanogaster development. Dev Cell. 2003;5:337–350. - PubMed
-
- Bernstein E, Caudy AA, Hammond SM, Hannon GJ. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature. 2001;409:363–366. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
