

Formation of monoterpenes in Antirrhinum majus and Clarkia breweri flowers involves heterodimeric geranyl diphosphate synthases
- PMID: 15031409
- PMCID: PMC412871
- DOI: 10.1105/tpc.020156
Formation of monoterpenes in Antirrhinum majus and Clarkia breweri flowers involves heterodimeric geranyl diphosphate synthases
Abstract
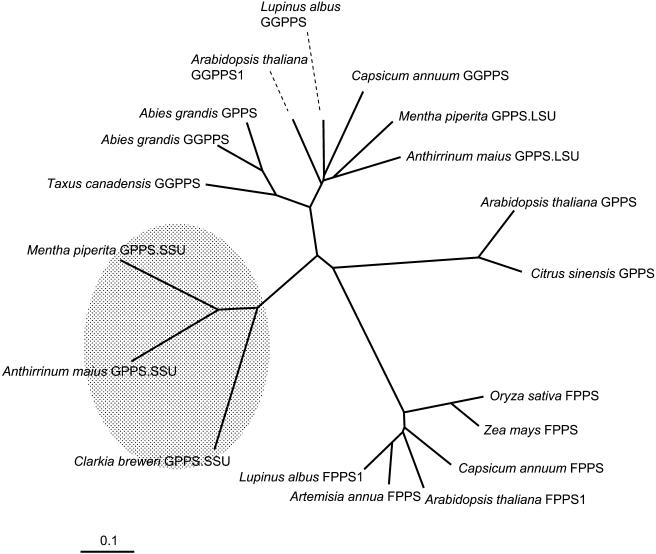
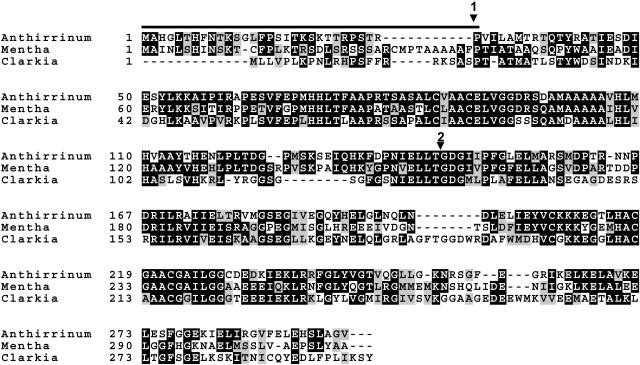
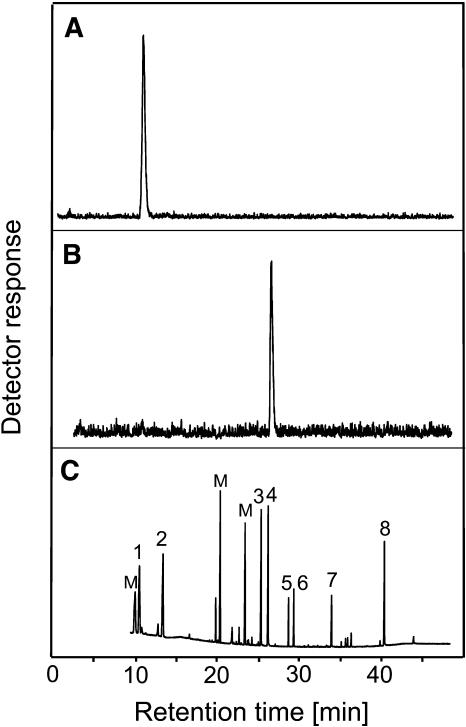
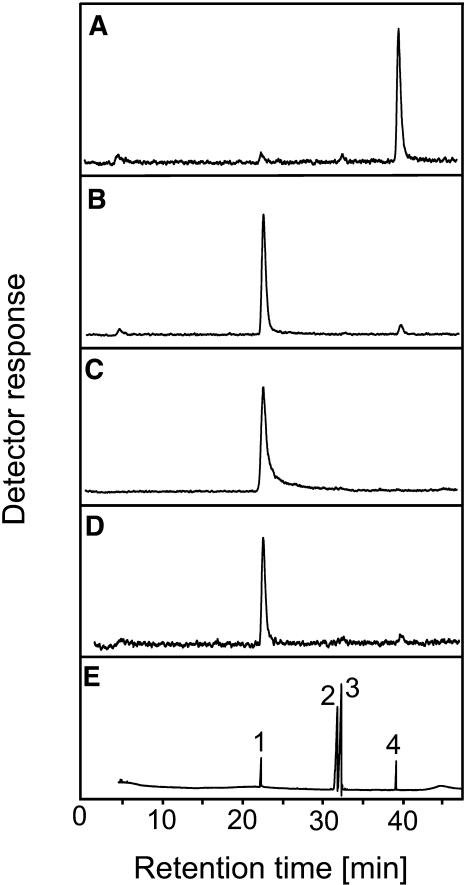
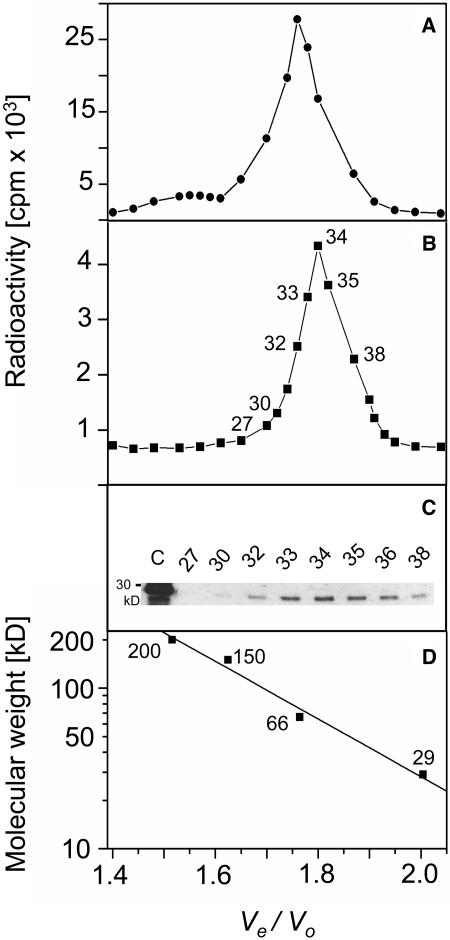
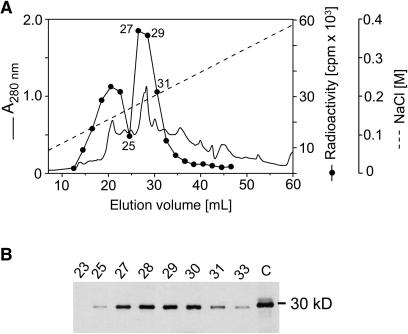
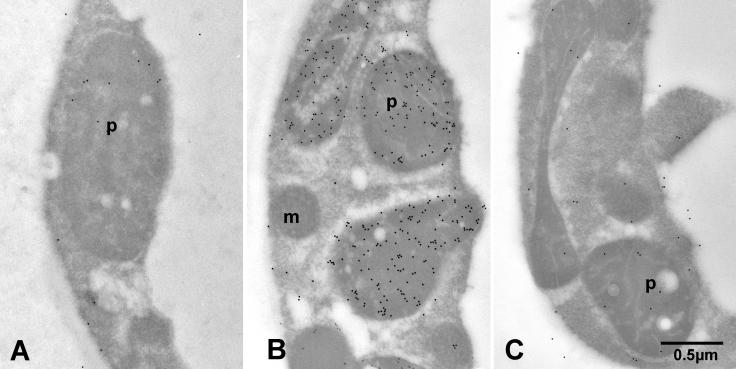
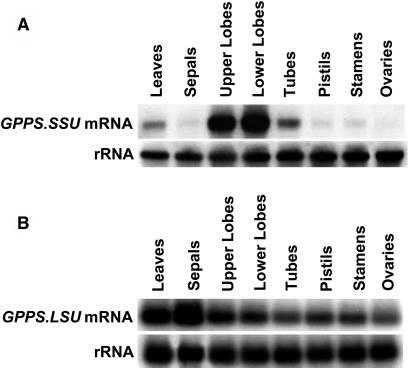
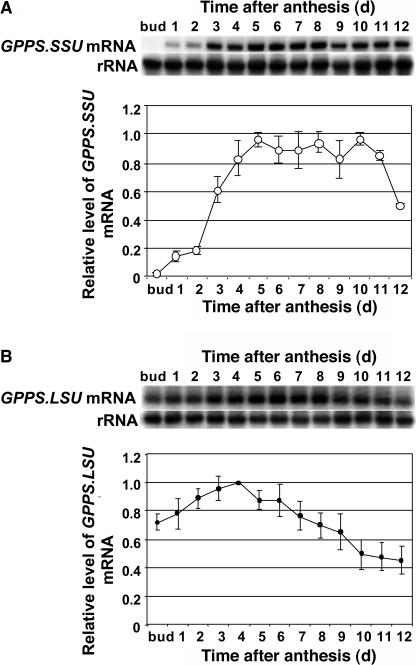
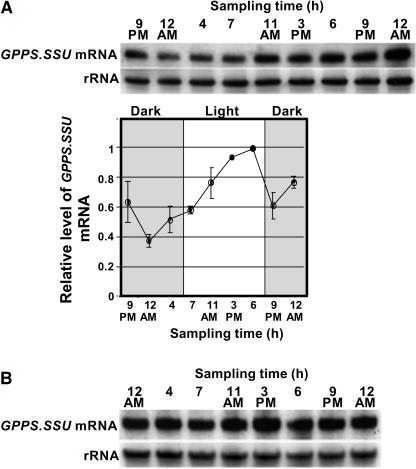
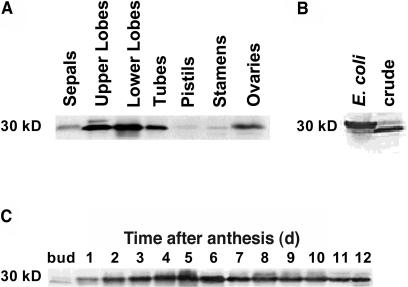
The precursor of all monoterpenes is the C10 acyclic intermediate geranyl diphosphate (GPP), which is formed from the C5 compounds isopentenyl diphosphate and dimethylallyl diphosphate by GPP synthase (GPPS). We have discovered that Antirrhinum majus (snapdragon) and Clarkia breweri, two species whose floral scent is rich in monoterpenes, both possess a heterodimeric GPPS like that previously reported from Mentha piperita (peppermint). The A. majus and C. breweri cDNAs encode proteins with 53% and 45% amino acid sequence identity, respectively, to the M. piperita GPPS small subunit (GPPS.SSU). Expression of these cDNAs in Escherichia coli yielded no detectable prenyltransferase activity. However, when each of these cDNAs was coexpressed with the M. piperita GPPS large subunit (GPPS.LSU), which shares functional motifs and a high level of amino acid sequence identity with geranylgeranyl diphosphate synthases (GGPPS), active GPPS was obtained. Using a homology-based cloning strategy, a GPPS.LSU cDNA also was isolated from A. majus. Its coexpression in E. coli with A. majus GPPS.SSU yielded a functional heterodimer that catalyzed the synthesis of GPP as a main product. The expression in E. coli of A. majus GPPS.LSU by itself yielded active GGPPS, indicating that in contrast with M. piperita GPPS.LSU, A. majus GPPS.LSU is a functional GGPPS on its own. Analyses of tissue-specific, developmental, and rhythmic changes in the mRNA and protein levels of GPPS.SSU in A. majus flowers revealed that these levels correlate closely with monoterpene emission, whereas GPPS.LSU mRNA levels did not, indicating that the levels of GPPS.SSU, but not GPPS.LSU, might play a key role in regulating the formation of GPPS and, thus, monoterpene biosynthesis.
Figures

References
-
- Bouvier, F., Suire, C., d'Harlingue, A., Backhaus, R.A., and Camara, B. (2000). Molecular cloning of geranyl diphosphate synthase and compartmentation of monoterpene synthesis in plant cells. Plant J. 24, 241–252. - PubMed
-
- Bradford, M.M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254. - PubMed
-
- Burke, C.C., and Croteau, R. (2002. a). Interactions with the small subunit of geranyl diphosphate synthase modifies the chain length specificity of geranylgeranyl diphosphate synthase to produce geranyl diphosphate. J. Biol. Chem. 277, 3141–3149. - PubMed
-
- Burke, C.C., and Croteau, R. (2002. b). Geranyl diphosphate synthase from Abies grandis: cDNA isolation, functional expression, and characterization. Arch. Biochem. Biophys. 405, 130–136. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
