

Ephrin-B1 forward and reverse signaling are required during mouse development
- PMID: 15037550
- PMCID: PMC374238
- DOI: 10.1101/gad.1171704
Ephrin-B1 forward and reverse signaling are required during mouse development
Abstract
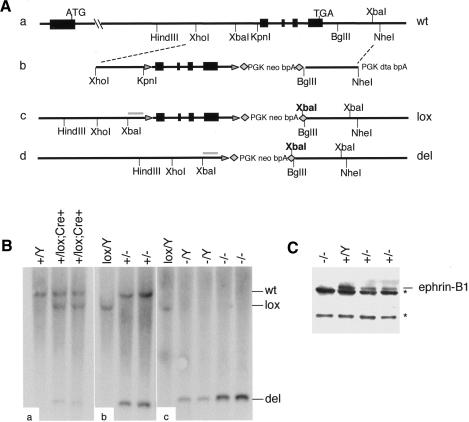
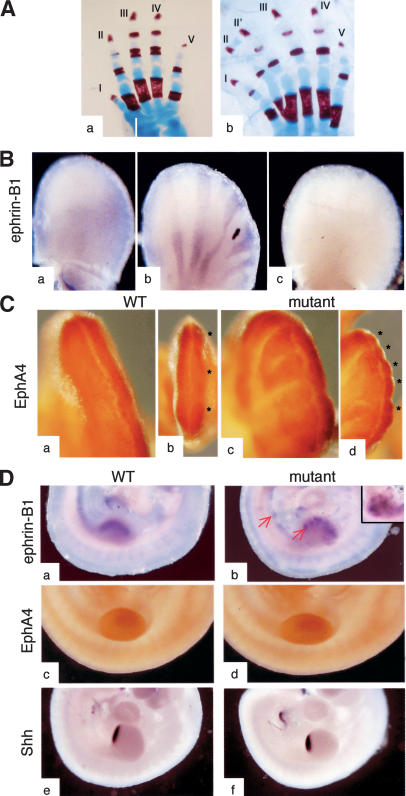
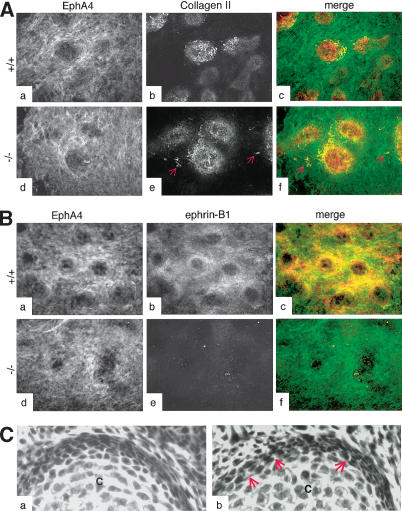
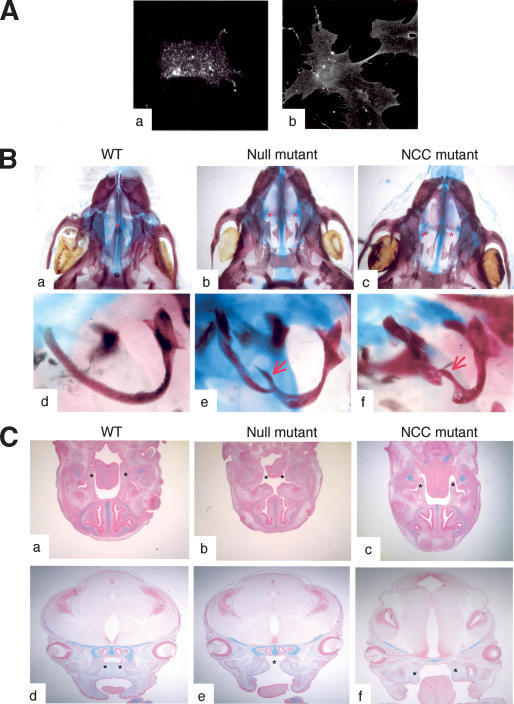
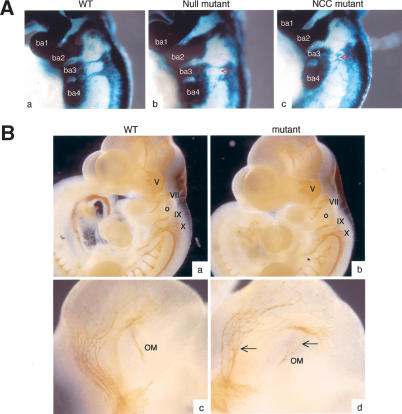
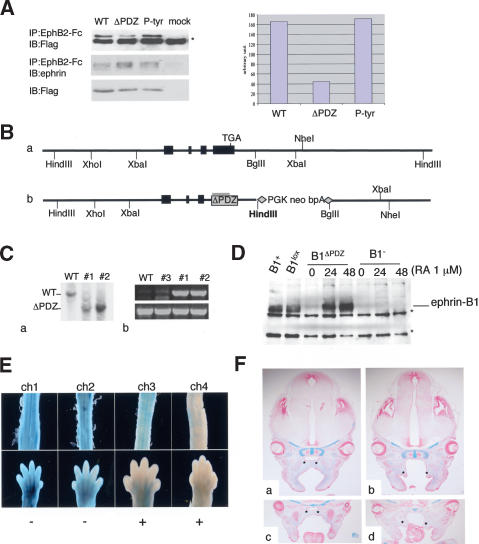
Eph receptors and ephrin ligands are key players in many developmental processes including embryo patterning, angiogenesis, and axon guidance. Eph/ephrin interactions lead to the generation of a bidirectional signal, in which both the Eph receptors and the ephrins activate downstream signaling cascades simultaneously. To understand the role of ephrin-B1 and the importance of ephrin-B1-induced reverse signaling during embryonic development, we have generated mouse lines carrying mutations in the efnb1 gene. Complete ablation of ephrin-B1 resulted in perinatal lethality associated with a range of phenotypes, including defects in neural crest cell (NCC)-derived tissues, incomplete body wall closure, and abnormal skeletal patterning. Conditional deletion of ephrin-B1 demonstrated that ephrin-B1 acts autonomously in NCCs, and controls their migration. Last, a mutation in the PDZ binding domain indicated that ephrin-B1-induced reverse signaling is required in NCCs. Our results demonstrate that ephrin-B1 acts both as a ligand and as a receptor in a tissue-specific manner during embryogenesis.
Figures
References
-
- Adams R.H. 2002. Vascular patterning by Eph receptor tyrosine kinases and ephrins. Semin. Cell Dev. Biol. 13: 55–60. - PubMed
-
- Adams R.H., Diella, F., Hennig, S., Helmbacher, F., Deutsch, U., and Klein, R. 2001. The cytoplasmic domain of the ligand ephrinB2 is required for vascular morphogenesis but not cranial neural crest migration. Cell 104: 57–69. - PubMed
-
- Bouillet P., Oulad-Abdelghani, M., Vacaire, S., Garnier, J.M., Schuhbaur, B., Dolle, P., and Chambon, P. 1995. Efficient cloning of cDNAs of retinoic acid-responsive genes in P19 embryonal carcinoma cells and characterization of a novel mouse gene, Stra1 (mouse LERK-2/Eplg2). Dev. Biol. 170: 420–433. - PubMed
-
- Bourgeois P., Bolcato-Bellemin, A.-L., Danse, J.-M., Bloch-Zupan, A., Yoshiba, K., Stoetzel, C., and Perrin-Schmitt, F. 1998. The variable expressivity and incomplete penetrance of the twist-null heterozygous mouse phenotype resemble those of human Saethre-Chotzen syndrome. Hum. Mol. Genet. 7: 945–957. - PubMed
-
- Bruckner K., Pasquale, E.B., and Klein, R. 1997. Tyrosine phosphorylation of transmembrane ligands for Eph receptors. Science 275: 1640–1643. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous