

Myosin-induced movement of alphaalpha, alphabeta, and betabeta smooth muscle tropomyosin on actin observed by multisite FRET
- PMID: 15041668
- PMCID: PMC1304079
- DOI: 10.1016/S0006-3495(04)74287-3
Myosin-induced movement of alphaalpha, alphabeta, and betabeta smooth muscle tropomyosin on actin observed by multisite FRET
Abstract
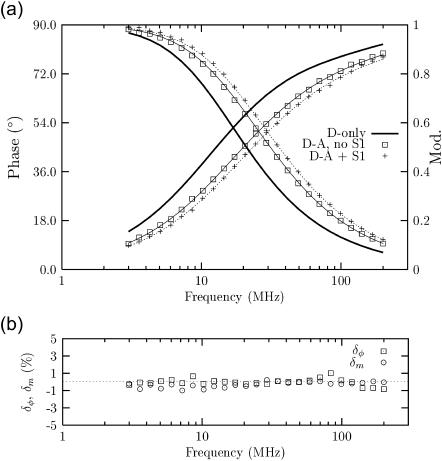
The interaction of the alphaalpha, betabeta, and alphabeta smooth muscle tropomyosin (Tm) isoforms with F-actin was systematically studied in the absence and in the presence of myosin subfragment 1 (S1) using multifrequency phase/modulation Förster resonance energy transfer (FRET). A Gaussian double distance distribution model was adopted to fit FRET data between a 5-(2-iodoacetyl-amino-ethyl-amino)naphthalene-1-sulfonic acid donor at either Cys-36 of the beta-chain or Cys-190 of the alpha-chain and a 4-dimethylaminophenylazophenyl 4'-maleimide acceptor at Cys-374 of F-actin. Experimental data were obtained for singly and doubly labeled alphabeta Tm (donor only at alpha, only at beta, or both) and for doubly labeled alphaalpha or betabeta Tm. Data for singly labeled alphabetaTm were combined in a global analysis with doubly labeled alphabetaTm. In all doubly labeled isoforms, upon S1 binding, one donor-acceptor "apparent" distance increased slightly by 0.5-2 A, whereas the other decreased by 6-9 A. These changes are consistent with a uniform "rolling" motion of Tm over the F-actin surface. The analysis indicates that Tm occupies relatively well-defined positions, with some flexibility, in both the predominantly closed (-S1) and open (+S1) thin-filament states. The results for the alphabetaTm heterodimer indicate that the local twofold symmetry of alphaalpha or betabeta Tm is effectively broken in alphabetaTm bound to F-actin, which implies a difference between the alpha- and beta-chains in terms of their interaction with F-actin.
Figures





References
-
- Arcioni, A., R. Tarroni, and C. Zannoni. 1993. Global target analysis of fluorescence depolarization in model membranes using exponential splines. J. Chem. Soc. Faraday Trans. 89:2815–2822.
-
- Bacchiocchi, C., M. Miki, T. Wakabayashi, and S. S. Lehrer. 2003. The C-terminal region of Tm does not exhibit Ca2+- induced movement in reconstituted thin filaments. Biophys. J. 84:214a. (Abstr.)
-
- Bernstein, H. J. 1999. RasMol 2.7.1. Molecular Graphics Visualization Tool (based on RasMol 2.6 by R. Sayle). http://www.bernstein-plus-sons.com/software/rasmol/.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
