Fig. 1.
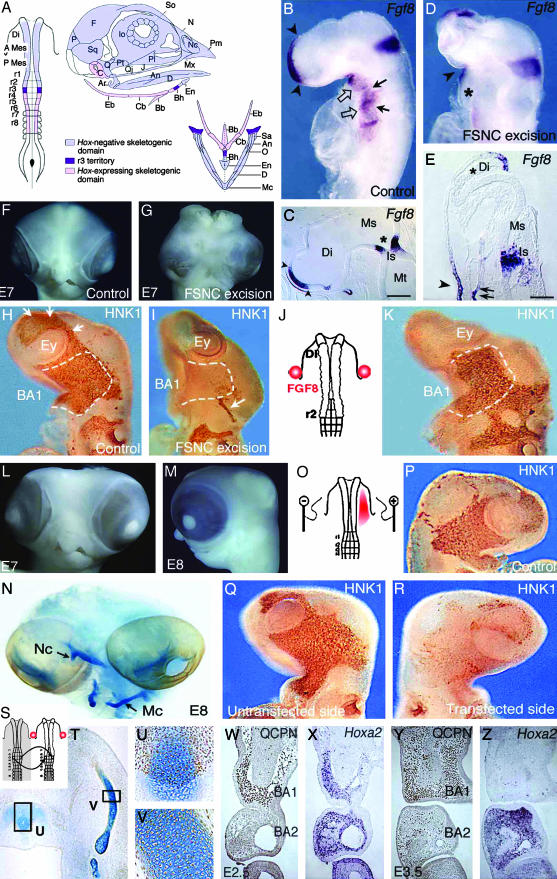
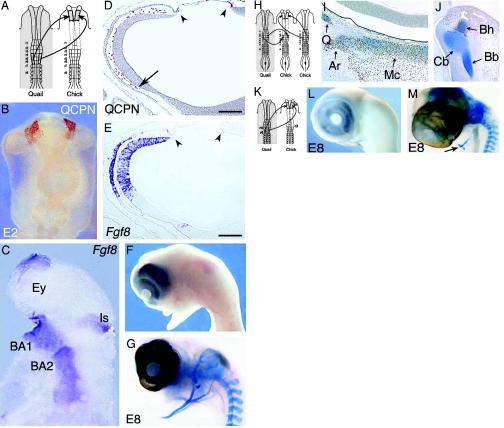
(On opposite page.) Influence of FSNC excision on Fgf8 expression in the embryonic head (A–E). (A) Fate map of the 5ss chicken embryo cephalic NF (2, 19). The anterior domain, extending from mid-diencephalon down through r2, yields Hox-negative NCC (in light blue); these NCC, which form the facial skeleton, are designated as FSNC. The posterior domain, extending from r4 through r8, generates Hox-positive NCC (in pink) and yields most of the hyoid cartilage. At the edge of both Hox-positive and Hox-negative domains, r3 NF is represented in violet. r3 NCC provide a minor contribution to both BA1 and BA2. Respective contributions of these domains to the craniofacial and hypobranchial skeleton are color-coded as described previously. (B) In situ hybridization with an Fgf8 riboprobe carried out on normal 24ss chick embryos in toto and (C) in sagittal section (C). The facial processes (nasal bud and BA1) are colonized by the FSNC. Fgf8 is expressed in the prosencephalon neuroepithelium and in the adjacent superficial ectoderm (arrowheads; B and C). A second site of Fgf8 expression is in the ectoderm of BA1, -2, and -3 through -4 (open arrows). First detected in the ventral ectoderm corresponding to the level of the presumptive BA1 from 13ss, Fgf8 expression becomes widely distributed at 24ss on the whole BA ectoderm with more intense foci of transcript accumulation facing the pharyngeal pouches (arrows). The third site of Fgf8 expression is in the neuroepithelium of the isthmus (B and C; *). In embryos in which resection of the FSNC is carried out bilaterally at 5–6ss from the mid-diencephalic level down to r3 (r3 excluded; D), striking changes are seen in Fgf8 expression during the 24 h after the resection. At 24ss, the prosencephalon is reduced in size, and the anterior Fgf8 expression site is less extended and displaced dorsally (D). On the sagittal section (E), FSNC extirpation affects the pattern of Fgf8 in the superficial ectoderm and precludes its expression in the prosencephalic neuroepithelium where transcript accumulation is completely abolished (*; E). The BAs are not properly developed, and expression of Fgf8 gene is very reduced (*) when compared to stage-matched controls (B and D). At the level of the isthmus, Fgf8 expression is less intense, more widespread, and essentially localized in the ventricular zone of the neural epithelium. Fgf8 expression is maintained in the stomodeal ectoderm (arrowhead on D and E) and in the endoderm of the foregut (double arrows on E). FSNC ablation completely abolishes the development of the face and brain [see E7 control (F) and operated (G) embryos]. Restoration of facial morphogenesis by exogenous FGF8 (H–N). Comparison of NC migration in control (H), FSNC-excised (I), and FSNC-excised subjected to exogenous FGF8 through beads placed at the presumptive level of BA1 ectoderm (J and K; HNK1 mAb staining at E3). In E3 control embryos, cephalic NCC, which appear as brown in toto, have spread laterally and massively populate the forming BA1 (dotted line). They encompass the optic vesicles and are in the process of migrating rostrally to colonize the nasofrontal bud (arrows). After FSNC excision, only rare HNK1-labeled cells are present in BA1. Note the labeling of the trigeminal ganglion (arrow) that attests to the contribution of midrhombencephalic NCC to the cranial peripheral nervous system. Treatment of FSNC-excised embryos with FGF8 rescues the colonization of BA1 by r3-derived cells. At E7 and E8, these embryos display a partial restoration of their facial morphology with the development of maxillary and mandibular components of the upper and lower beaks, respectively (L and M). (N) Alcian blue staining of the skeletal pieces reveals a reduced nasal capsule (Nc) in the upper face and the entire set of lower jaw cartilages among which Meckel's (Mc) is recognizable. Inhibition of RNA processing by RNAi using Fgf8-dsRNA (O–R). In normal chick embryos, Fgf8-dsRNA has been unilaterally electroporated in the BA1 presumptive ectoderm (red area; O). At E3 on both control (P) and experimental (Q, untransfected side) embryos, NCC fully populate the maxillo-mandibular processes. In contrast, on the transfected side (R), no (or very little) colonization of NCC occurs, suggesting a role of the FGF8 protein as an attractant for NCC migration. Maxillo-mandibular regeneration originates from r3-derived NCC (S–Z). After the bilateral excision of the FSNC, the r3 NF of the host embryo was bilaterally replaced by its quail counterpart and FGF8 beads were supplied as before (S). Quail cell detection using QCPN mAb combined with the Alcian blue counterstaining to evidence cartilaginous structures shows that r3-derived crest cells are the exclusive source of the chondroblasts that generate the anterior part of basihyal (T and U) and Meckel's cartilage (T and V) in the lower jaw. Note that the posterior part of basihyal is normally of r4 origin (2, 19). In this context, at E2.5, the r3-derived NCC that are in the process of populating the BA1 (W) express Hoxa2 (X). At E3.5, they turn to an Hox-negative status (Y and Z), although those that are in BA2 exhibit accumulation of the Hoxa2 transcript. An, angular; Ar, articular; A Mes, anterior mesencephalon; BA, branchial arch; Bb, basibranchial; Bh, basihyal; C, columella; Cb, ceratobranchial; D, dentary; Di, diencephalon; Eb, epibranchial; En, entoglossum; Ey, eye; F, frontal; Io, interorbital septum; Is, isthmus; J, jugal; Ms, mesencephalon; Mt, metencephalon; Mx, maxillary; N, nasal; Nc, nasal capsule; O, opercular; P, parietal; Pl, palate; Pt, pterygoid; P mes, posterior mesencephalon; Q, quadrate; Qj, quadratojugal; r, rhombomere; Sa, supraangular; So, sclerotic ossicles; Sq, squamosal. [Scale bars: 350 μm(C) and 100 μm(E).]