

Review
doi: 10.1038/nrmicro817.
Virus entry: molecular mechanisms and biomedical applications
Affiliations
- PMID: 15043007
- PMCID: PMC7097642
- DOI: 10.1038/nrmicro817
Item in Clipboard
Review
Virus entry: molecular mechanisms and biomedical applications
Nat Rev Microbiol.
2004 Feb.
Abstract
Viruses have evolved to enter cells from all three domains of life--Bacteria, Archaea and Eukaryotes. Of more than 3,600 known viruses, hundreds can infect human cells and most of those are associated with disease. To gain access to the cell interior, animal viruses attach to host-cell receptors. Advances in our understanding of how viral entry proteins interact with their host-cell receptors and undergo conformational changes that lead to entry offer unprecedented opportunities for the development of novel therapeutics and vaccines.
Conflict of interest statement
The author declares no competing financial interests.
Figures
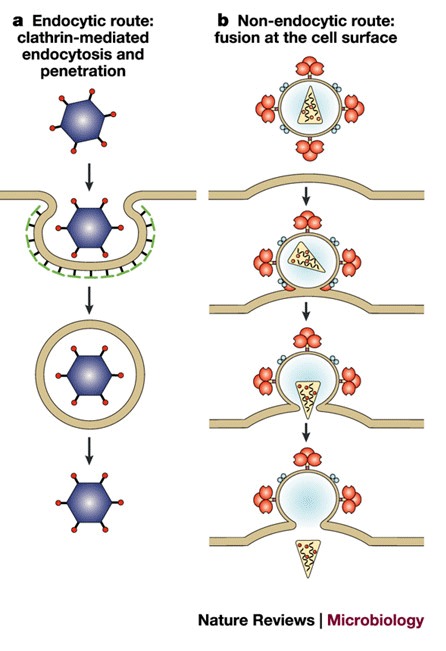
a | Clathrin-mediated endocytosis, for example, adenovirus. Endocytosis by caveolae can also occur, for example, SV40. b | Fusion at the cell membrane, for example, HIV.Fusion can also occur from inside an endosome, for example, influenza.
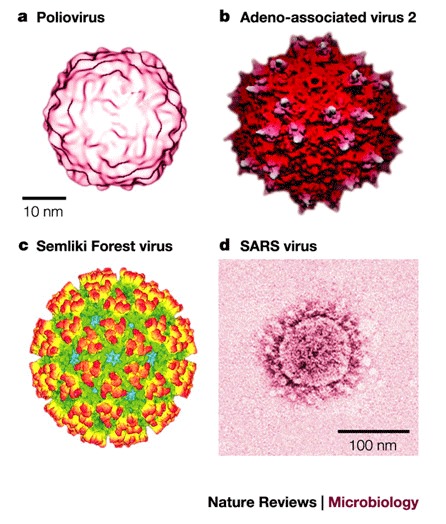
a | Structure of the 160S poliovirus particle. Reproduced with permission from Ref. © (2000) American Society for Microbiology. b | Surface topology of adeno-associated virus 2. The protruding spikes are coloured white. Reproduced with permission from Ref. © (2002) National Academies of Sciences, USA. c | Structure of Semliki Forest virus. The colour scheme reflects the radial distance from the centre of the virion, increasing from blue to red. Reproduced with permission from Ref. © (2000) Elsevier Science. d | Structure of the severe acute respiratory syndrome (SARS) coronavirus. The coronavirus particle has club-shaped surface projections — known as spikes. Adeno-associated virus 2 and Semliki Forest virus particles (not to scale) are comparable in size to Poliovirus. Reproduced with permission from Ref. © (2003) Massachusetts Medical Society.
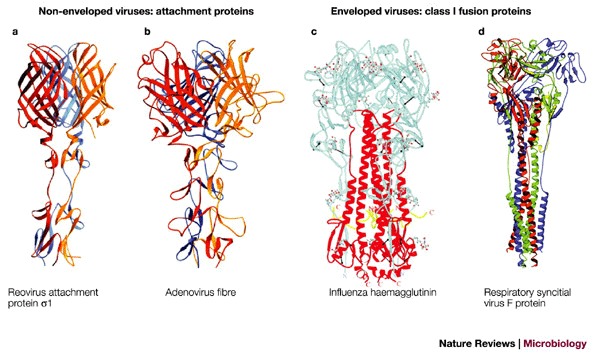
a | Ribbon tracing of reovirus attachment protein σ1. Reproduced with permission from Ref. © (2003) Wiley. b | Ribbon tracing of adenovirus fibre. Reproduced with permission from Ref. © (1985) American Association of Sciences. Both reovirus attachment protein σ1 (a) and adenovirus fibre (b) are homotrimers. The three monomers in each trimer are shown in red, orange and blue. Both proteins have head-and-tail morphology, with a triple β–spiral domain forming the tail and an eight-stranded-sandwich domain forming the head. c | Cleaved influenza haemagglutinin trimer. Reproduced with permission from Ref. © (2000) Annual Reviews. d | Model of the respiratory syncitial virus F protein structure (RSV-F), which is based on amino-acid sequence homology with the structure of the Newcastle disease virus F protein. Reproduced with permission from Ref. © (2003) Elsevier Science.
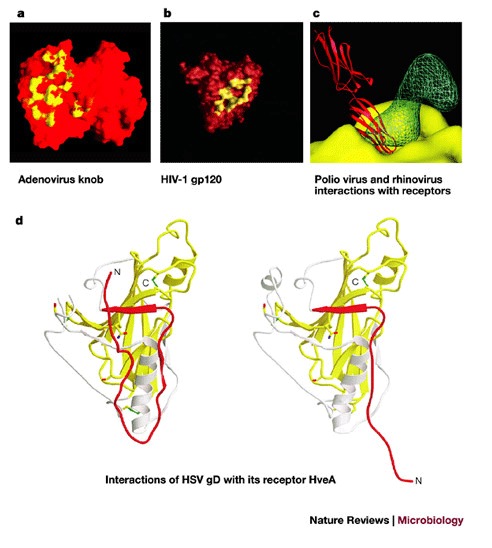
a | A molecular-surface representation of the interface between the adenovirus 12 (Ad12) knob and the coxsackievirus-adenovirus receptor D1 (CAR D1). The figure shows two adjacent Ad12 knob monomers, viewed at the interface between the Ad12 and CAR D1 molecules and coloured on a scale from yellow (contact with CAR D1) to red (no contact with CAR D1). Atoms in contact with CAR D1 are shared between the two Ad12 monomers. From Ref. © (1999) American Association of Sciences. b | The HIV-1 gp120 and CD4 receptor contact surface. The gp120 surface is shown in red, with the surface that is 3.5 Å distant from the CD4 receptor (surface-to-atom-centre distance) shown in yellow. From Ref. © (1998) Nature, Macmillan Magazines. c | Interaction of rhinovirus and poliovirus with their receptors. Comparison of poliovirus binding to its receptor CD155 and rhinovirus binding to its receptor ICAM-1. The figure shows the electron density of a poliovirus-bound CD155 molecule (green), the poliovirus molecular surface (yellow), including the canyon, and a rhinovirus-bound IC1 molecule (red). Structures of poliovirus and rhinovirus were superimposed to generate the figure. Reproduced with permission from Ref. © (2000) Oxford University Press. d | The conformational change of soluble glycoprotein D (gD285) fragment. The left panel shows the gD285 fragment, in complex with herpesvirus entry mediator A (HveA). The amino-terminal portion of the HveA binding hairpin (red) can be seen interacting with residues 224–240 of an α-helix of gD285 (white). The right panel shows unliganded gD285. From Ref. © (2003) American Society for Microbiology.
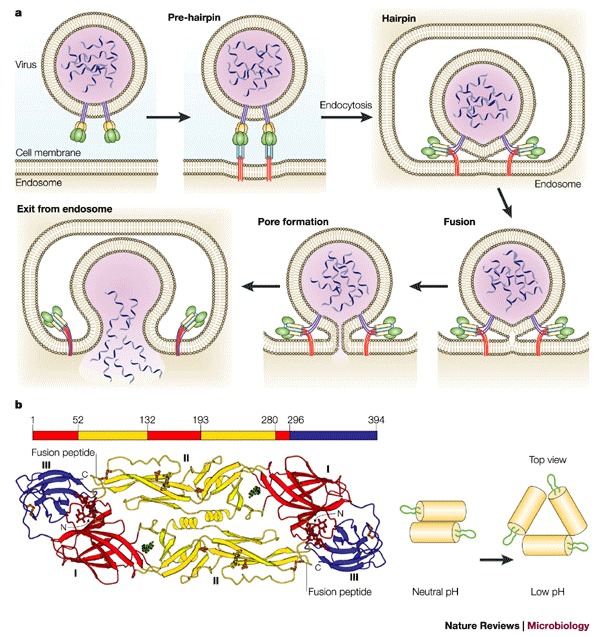
a | Schematic representation of a working model for viral membrane fusion mediated by class I fusion proteins. Influenza virus, which is internalized into an endosome, is shown as an example. In the native state of the fusion protein — which is a trimer — most of the surface subunit (green) is exposed. Part of the transmembrane subunit, including the fusion peptide, is not exposed. Following fusion-activating conditions, conformational changes occur to 'free' the fusion peptide (red) from its previously unexposed location. In the case of influenza HA, this occurs by a 'spring-loaded' mechanism. The 'pre-hairpin' intermediate spans two membranes — with the transmembrane domain positioned in the viral membrane and the fusion peptide inserted into the host-cell membrane. The pre-hairpin intermediate forms a trimer of hairpins, and membrane fusion occurs, which leads to pore formation and release of the viral genome into the cytoplasm. Modified with permission from Ref. © (2001) Annual Reviews. b | Conformational changes of class II viral fusion proteins and entry. Structure of fragments of the class II fusion (E) glycoprotein from dengue virus. The polypeptide chain begins in the central domain (domain I, red), which is an eight-stranded β-barrel with up-and-down topology. Two long insertions between strands in the central domain form the dimerization domain (domain II, yellow). The carboxy-terminal domain (domain III, blue) is an antiparallel β-barrel with an immunoglobulin-like topology, which is stabilized by three disulphide bridges. The domain definition is also highlighted on the peptide sequence (top). During entry, the oligomeric structure is reorganized. The configuration of dengue virus glycoproteins on the virion surface at neutral pH and the proposed configuration at low pH are shown. The E glycoproteins are shown as yellow cylinders and the fusion peptide is green. Reproduced from Refs , © (2002) Elsevier; (2003) National Academies of Sciences.
References
-
- d'Herelle F. The Bacteriophage and its Behavior. 1926.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
