

Protein kinase A activation promotes plasma membrane insertion of DCC from an intracellular pool: A novel mechanism regulating commissural axon extension
- PMID: 15044543
- PMCID: PMC6729852
- DOI: 10.1523/JNEUROSCI.4934-03.2004
Protein kinase A activation promotes plasma membrane insertion of DCC from an intracellular pool: A novel mechanism regulating commissural axon extension
Abstract
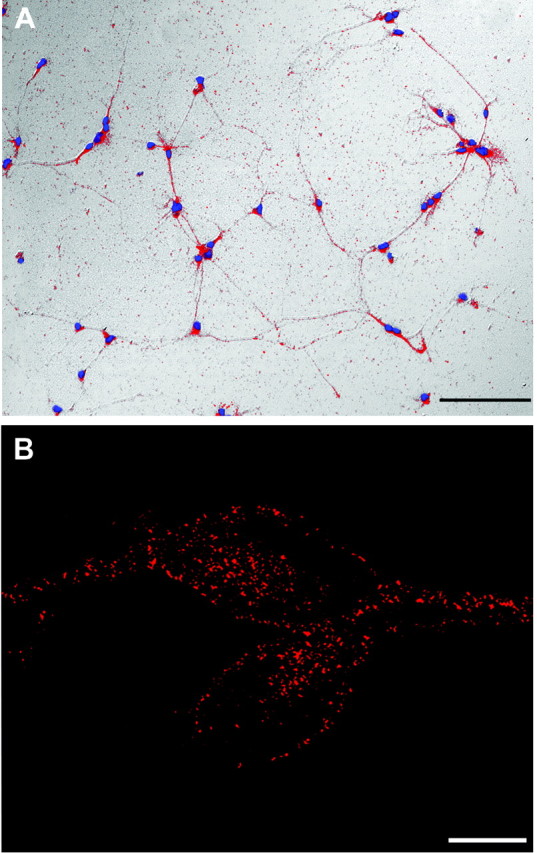
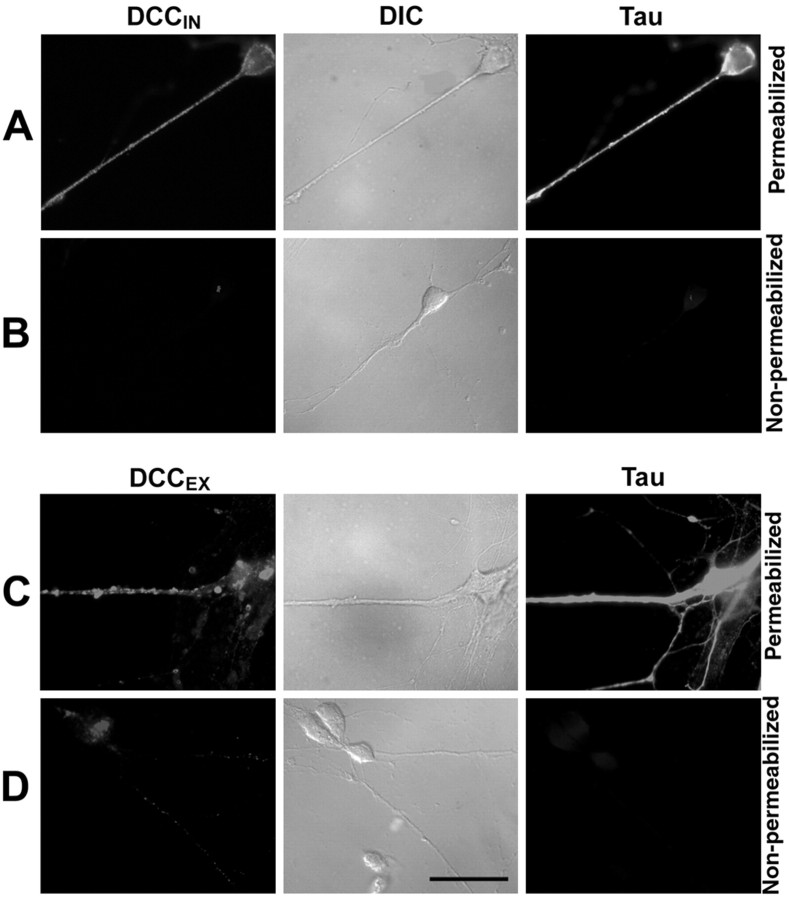
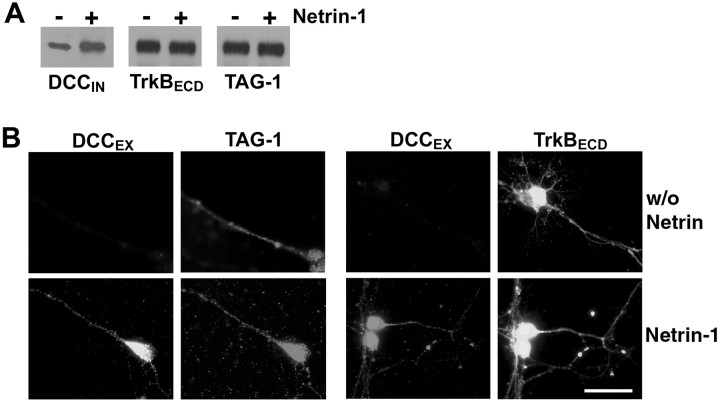
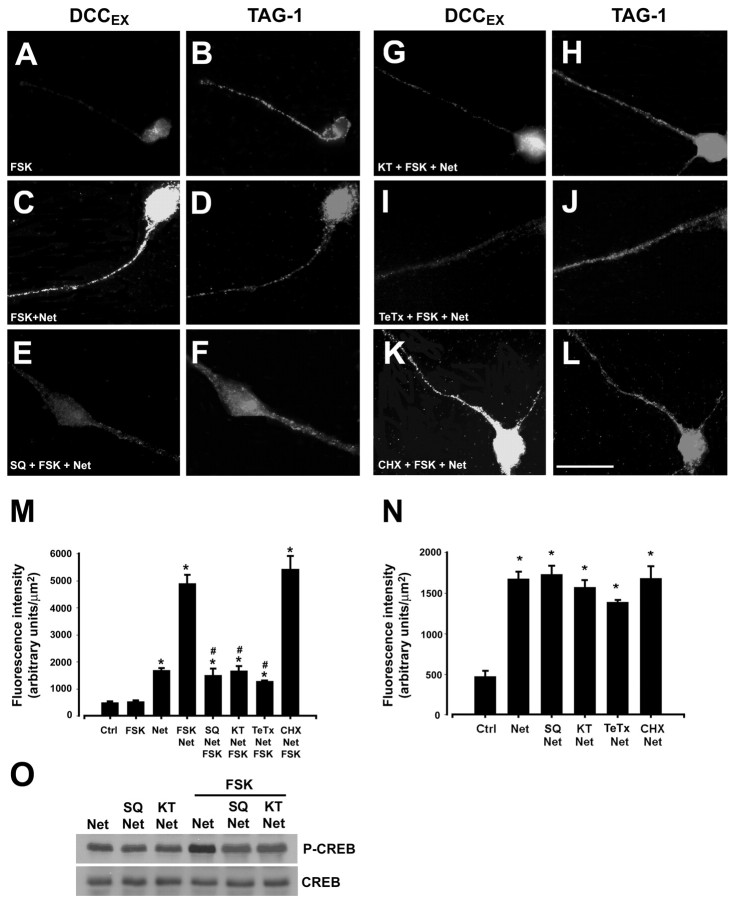
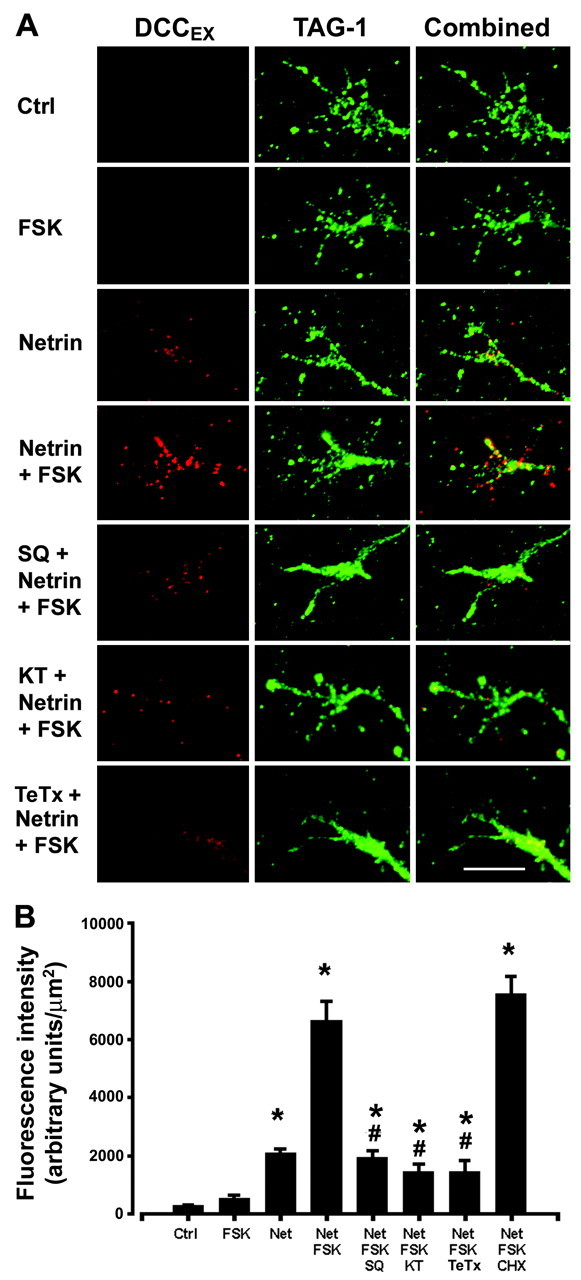
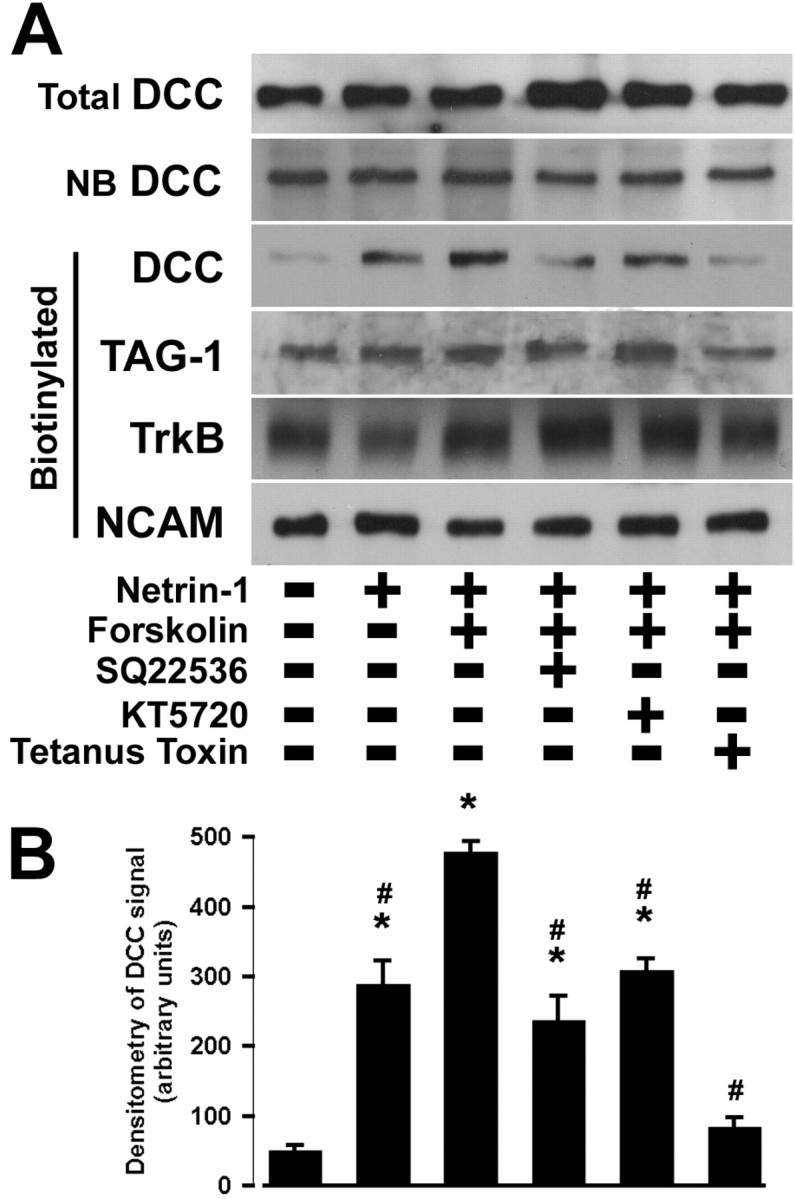
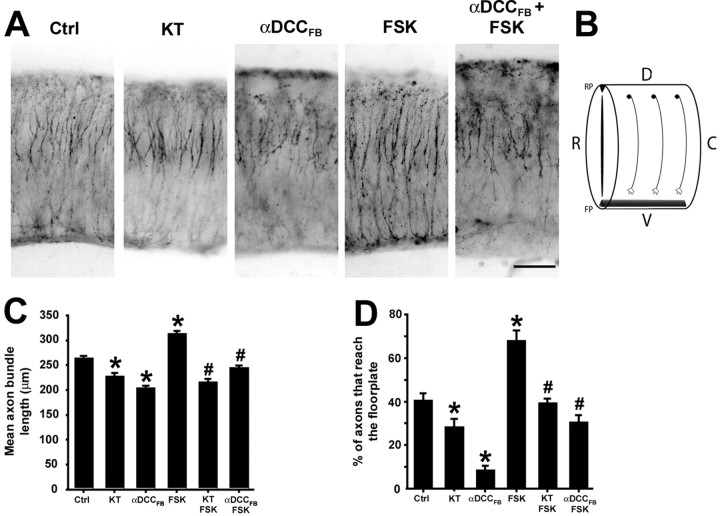
Protein kinase A (PKA) exerts a profound influence on axon extension during development and regeneration; however, the molecular mechanisms underlying these effects of PKA are not understood. Here, we show that DCC (deleted in colorectal cancer), a receptor for the axon guidance cue netrin-1, is distributed both at the plasma membrane and in a pre-existing intracellular vesicular pool in embryonic rat spinal commissural neurons. We hypothesized that the intracellular pool of DCC could be mobilized to the plasma membrane and enhance the response to netrin-1. Consistent with this, we show that application of netrin-1 causes a modest increase in cell surface DCC, without increasing the intracellular concentration of cAMP or activating PKA. Intriguingly, activation of PKA enhances the effect of netrin-1 on DCC mobilization and increases axon extension in response to netrin-1. PKA-dependent mobilization of DCC to the plasma membrane is selective, because the distributions of transient axonal glycoprotein-1, neural cell adhesion molecule, and trkB are not altered by PKA in these cells. Inhibiting adenylate cyclase, PKA, or exocytosis blocks DCC translocation on PKA activation. These findings indicate that netrin-1 increases the amount of cell surface DCC, that PKA potentiates the mobilization of DCC to the neuronal plasma membrane from an intracellular vesicular store, and that translocation of DCC to the cell surface increases axon outgrowth in response to netrin-1.
Figures


References
-
- Ackerman SL, Kozak LP, Przyborski SA, Rund LA, Boyer BB, Knowles BB (1997) The mouse rostral cerebellar malformation gene encodes an UNC-5-like protein. Nature 386: 838–842. - PubMed
-
- Barker PA, Shooter EM (1994) Disruption of NGF binding to the low affinity neurotrophin receptor p75LNTR reduces NGF binding to TrkA on PC12 cells. Neuron 13: 203–215. - PubMed
-
- Brittis PA, Lu Q, Flanagan JG (2002) Axonal protein synthesis provides a mechanism for localized regulation at an intermediate target. Cell 110: 223–235. - PubMed
-
- Brummendorf T, Lemmon V (2001) Immunoglobulin superfamily receptors: cis-interactions, intracellular adapters and alternative splicing regulate adhesion. Curr Opin Cell Biol 13: 611–618. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources