

Prevention of autoimmunity by targeting a distinct, noninvariant CD1d-reactive T cell population reactive to sulfatide
- PMID: 15051763
- PMCID: PMC2211873
- DOI: 10.1084/jem.20031389
Prevention of autoimmunity by targeting a distinct, noninvariant CD1d-reactive T cell population reactive to sulfatide
Abstract
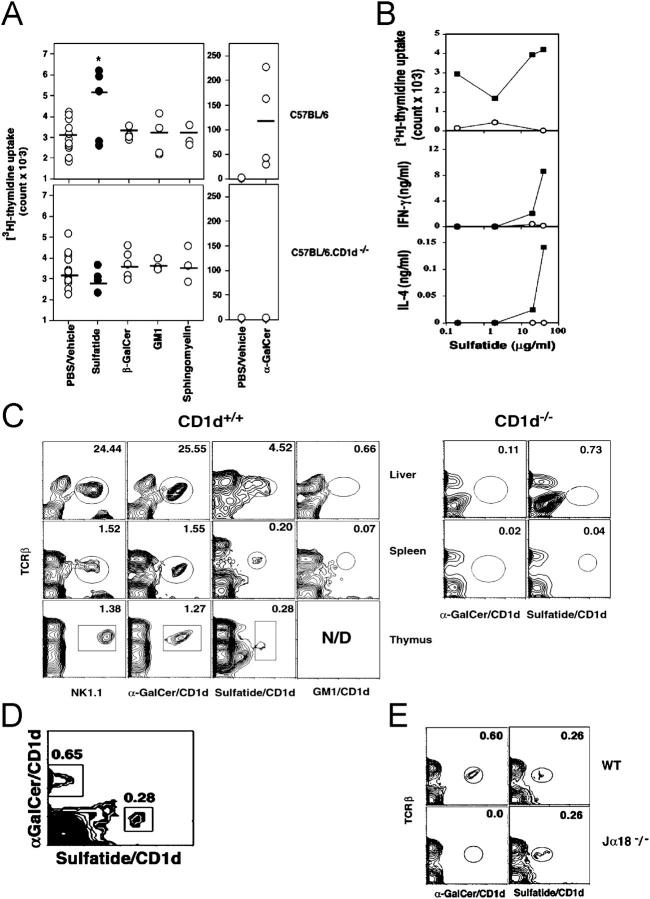
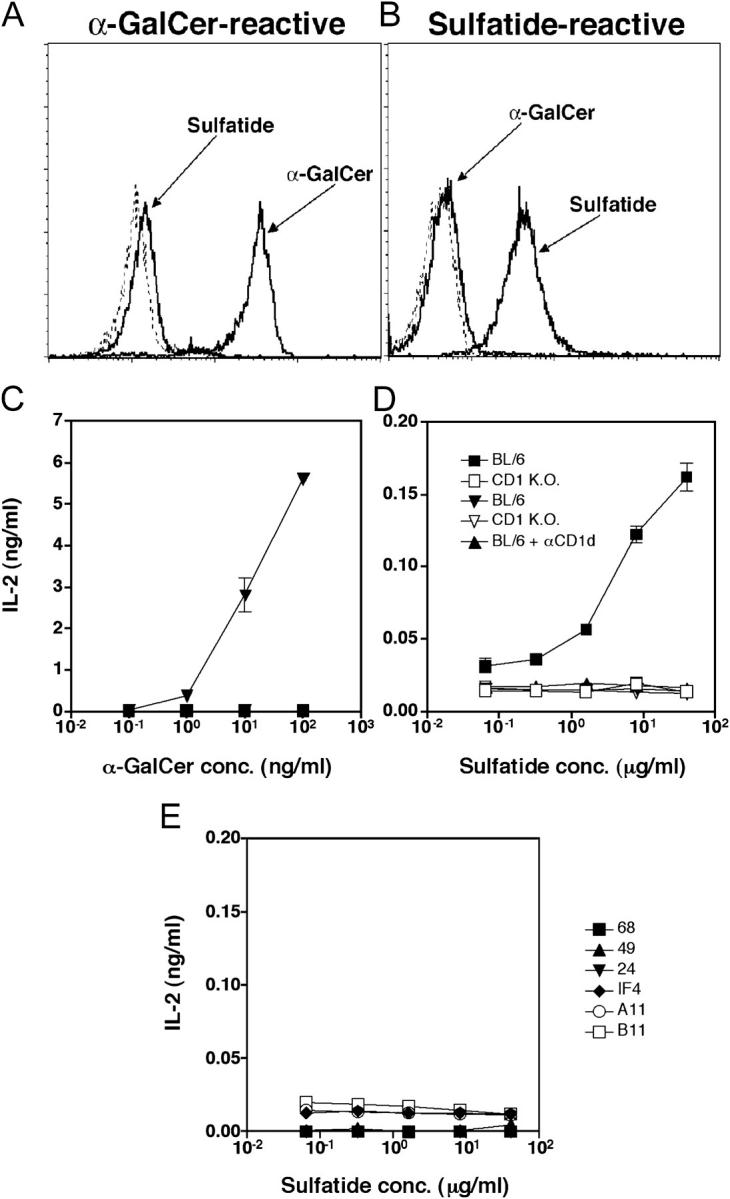
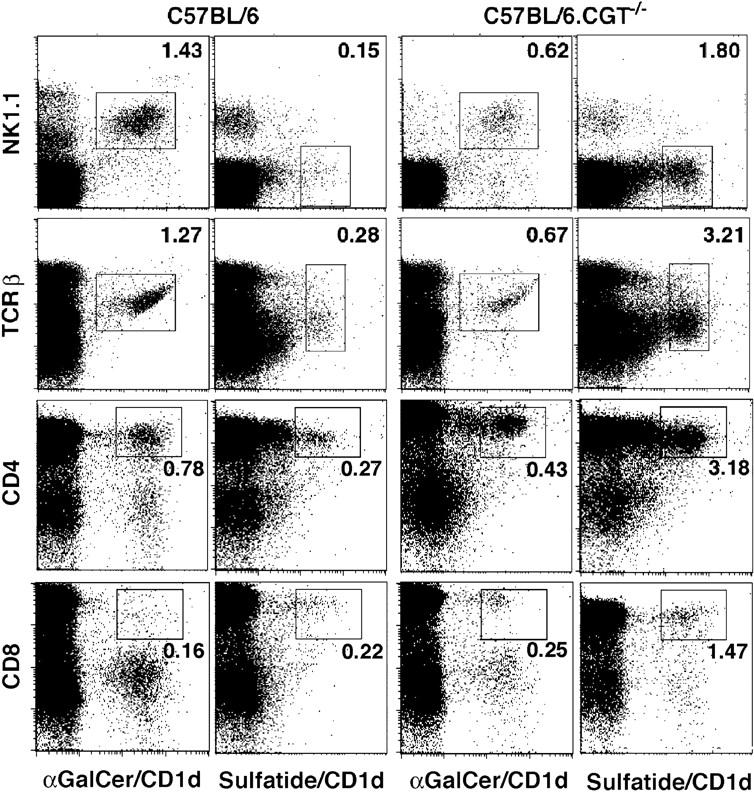
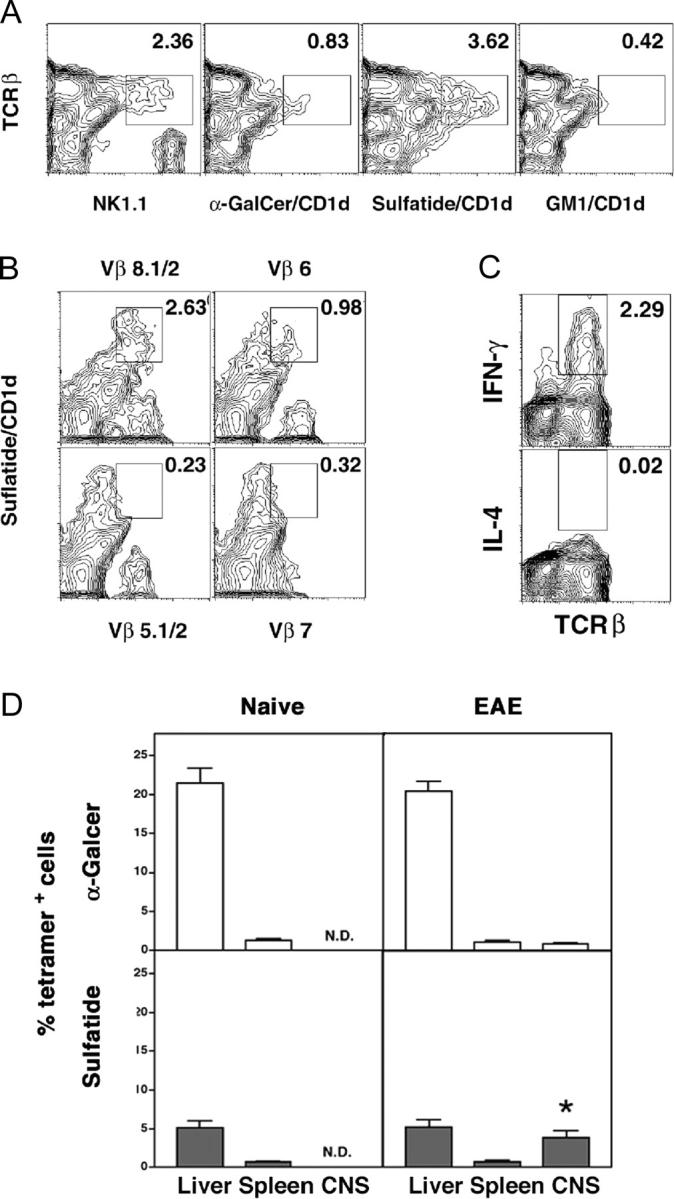
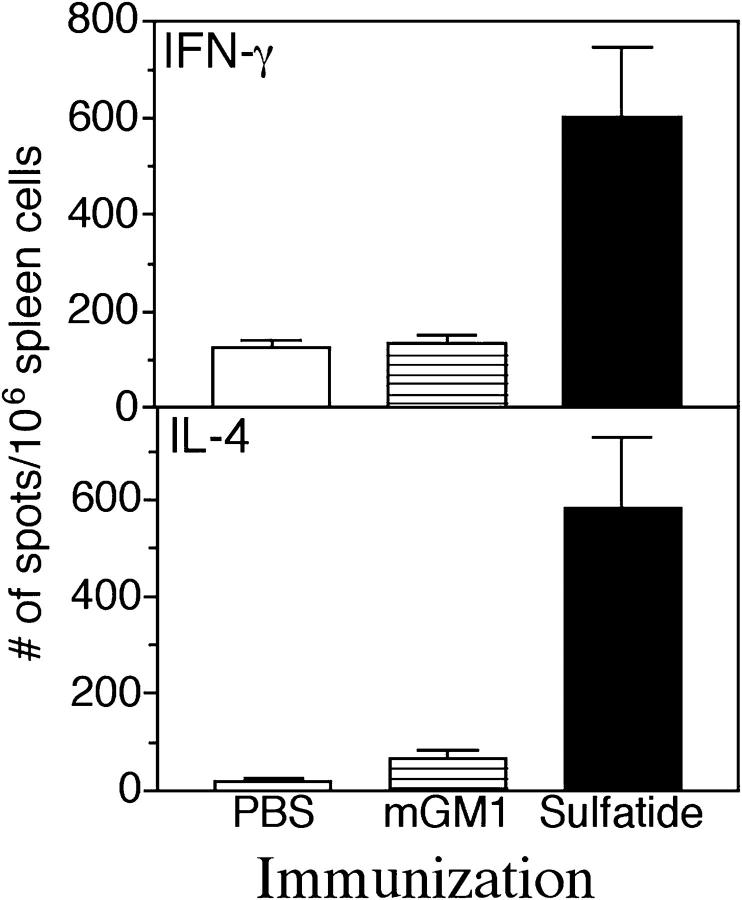
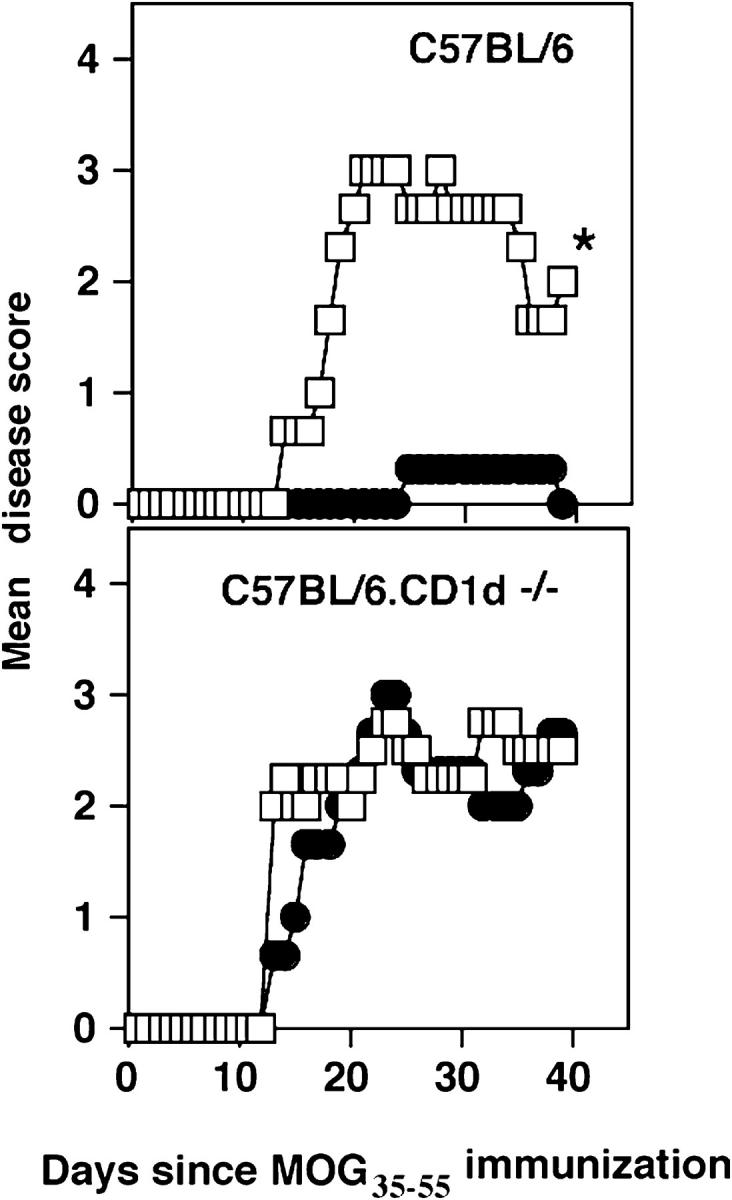
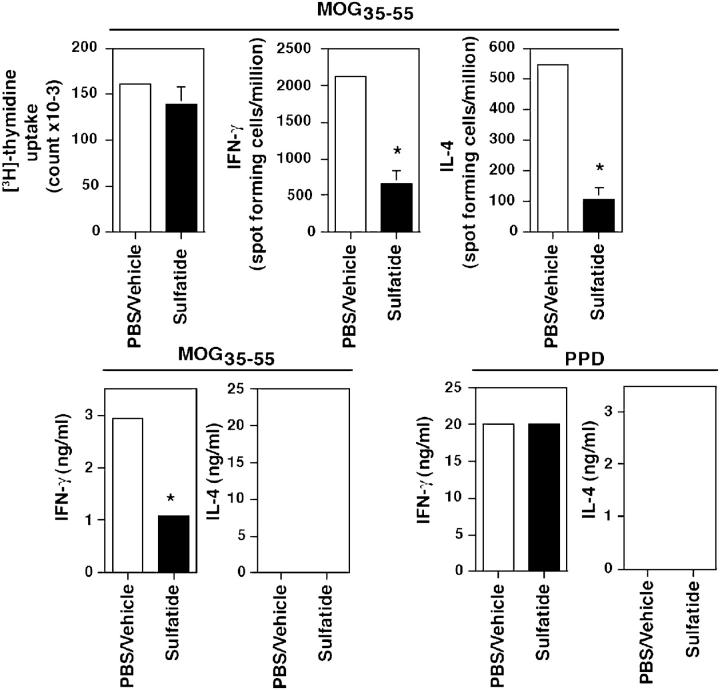
Class I and class II MHC-restricted T cells specific for proteins present in myelin have been shown to be involved in autoimmunity in the central nervous system (CNS). It is not yet known whether CD1d-restricted T cells reactive to myelin-derived lipids are present in the CNS and might be targeted to influence the course of autoimmune demyelination. Using specific glycolipid-CD1d tetramers and cloned T cells we have characterized a T cell population reactive to a myelin-derived glycolipid, sulfatide, presented by CD1d. This population is distinct from the invariant Valpha14+ NK T cells, and a panel of Valpha3/Valpha8+ CD1d-restricted NK T cell hybridomas is unable to recognize sulfatide in the presence of CD1d+ antigen-presenting cells. Interestingly, during experimental autoimmune encephalomyelitis a model for human multiple sclerosis, sulfatide-reactive T cells but not invariant NK T cells are increased severalfold in CNS tissue. Moreover, treatment of mice with sulfatide prevents antigen-induced experimental autoimmune encephalomyelitis in wild-type but not in CD1d-deficient mice. Disease prevention correlates with the ability of sulfatide to suppress both interferon-gamma and interleukin-4 production by pathogenic myelin oligodendrocyte glycoprotein-reactive T cells. Since recognition of sulfatide by CD1d-restricted T cells has now been shown both in mice and humans, study of murine myelin lipid-reactive T cells may form a basis for the development of intervention strategies in human autoimmune demyelinating diseases.
Figures
References
-
- Porcelli, S., C.T. Morita, and M.B. Brenner. 1992. CD1b restricts the response of human CD4-8− T lymphocytes to a microbial antigen. Nature. 360:593–597. - PubMed
-
- Sieling, P.A., D. Chatterjee, S.A. Porcelli, T.I. Prigozy, R.J. Mazzaccaro, T. Soriano, B.R. Bloom, M.B. Brenner, M. Kronenberg, P.J. Brennan, et al. 1995. CD1-restricted T cell recognition of microbial lipoglycan antigens. Science. 269:227–230. - PubMed
-
- Bendelac, A., M.N. Rivera, S.H. Park, and J.H. Roark. 1997. Mouse CD1-specific NK1 T cells: development, specificity, and function. Annu. Rev. Immunol. 15:535–562. - PubMed
-
- Porcelli, S.A., B.W. Segelke, M. Sugita, I.A. Wilson, and M.B. Brenner. 1998. The CD1 family of lipid antigen-presenting molecules. Immunol. Today. 19:362–368. - PubMed
-
- Moody, D.B., and S.A. Porcelli. 2003. Intracellular pathways of CD1 antigen presentation. Nat. Rev. Immunol. 3:11–22. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
