

A defect in nucleosome remodeling prevents IL-12(p35) gene transcription in neonatal dendritic cells
- PMID: 15051764
- PMCID: PMC2211877
- DOI: 10.1084/jem.20031272
A defect in nucleosome remodeling prevents IL-12(p35) gene transcription in neonatal dendritic cells
Abstract
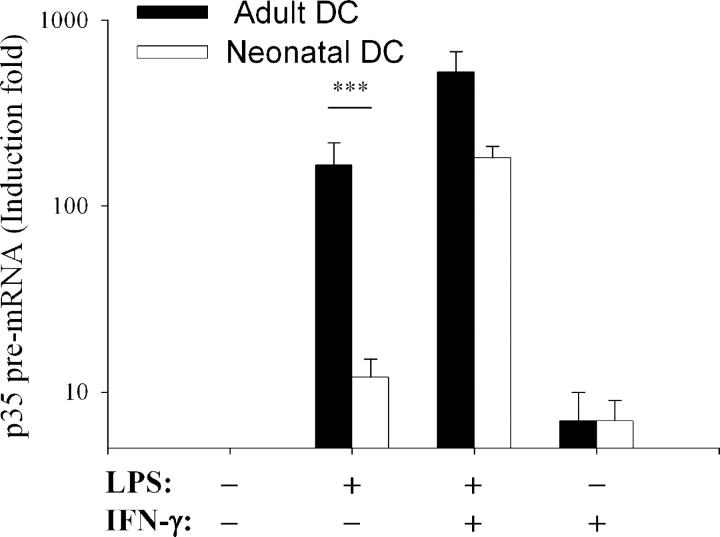
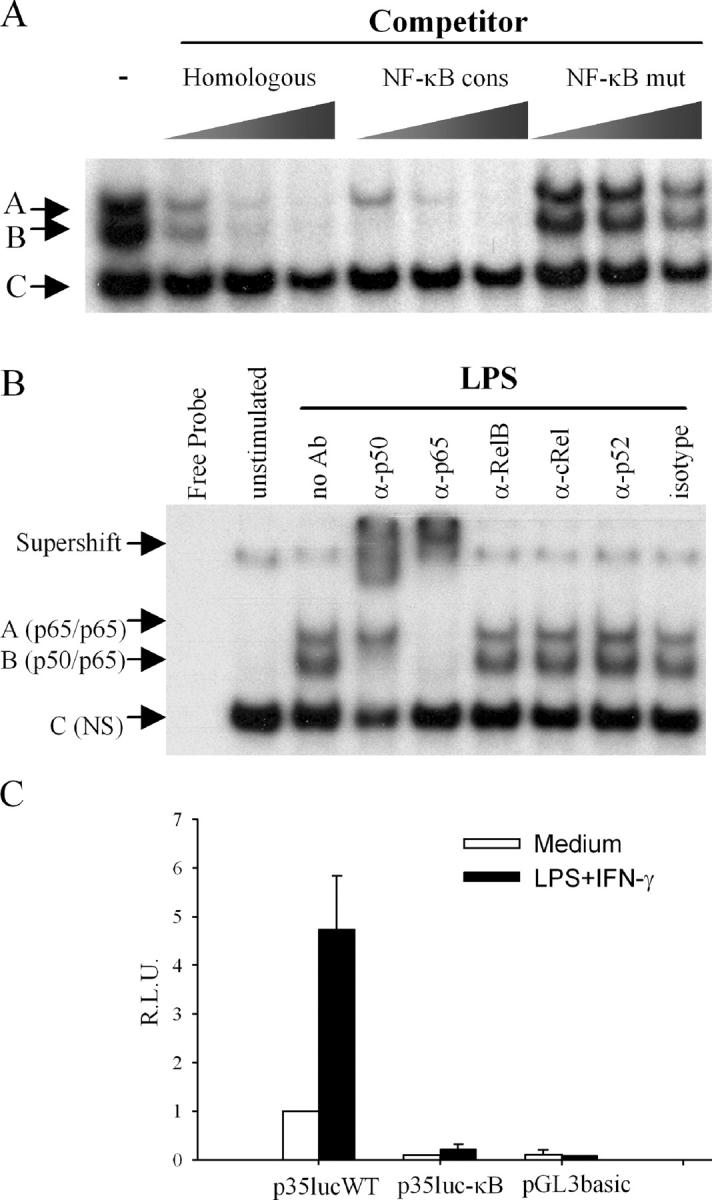
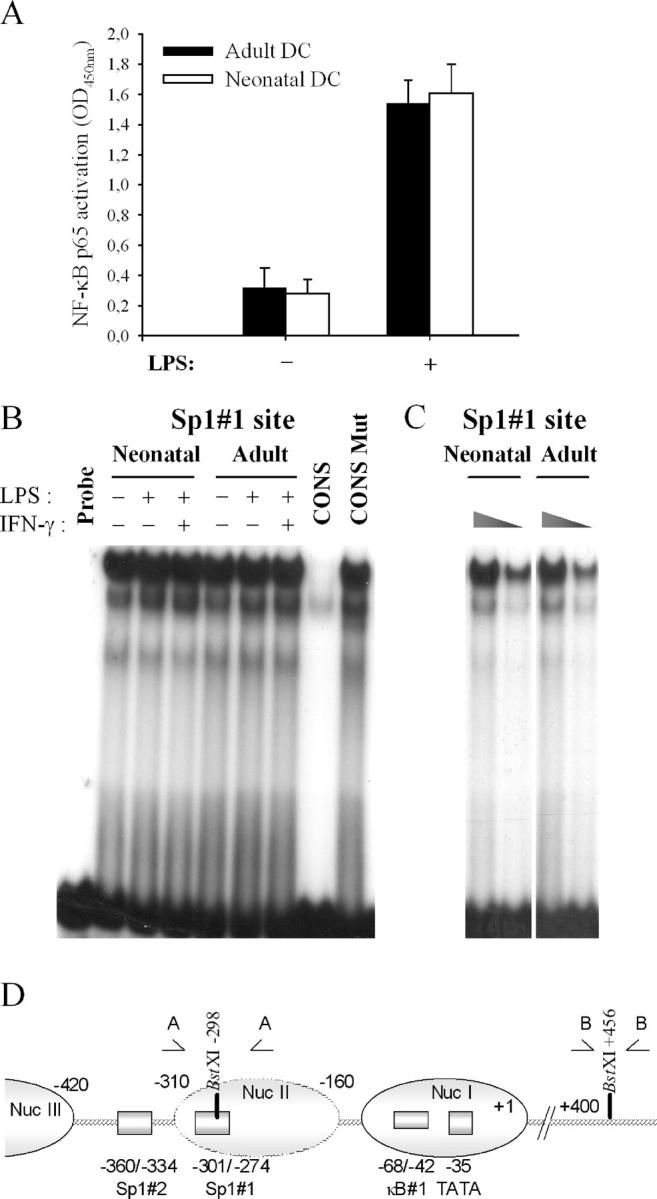
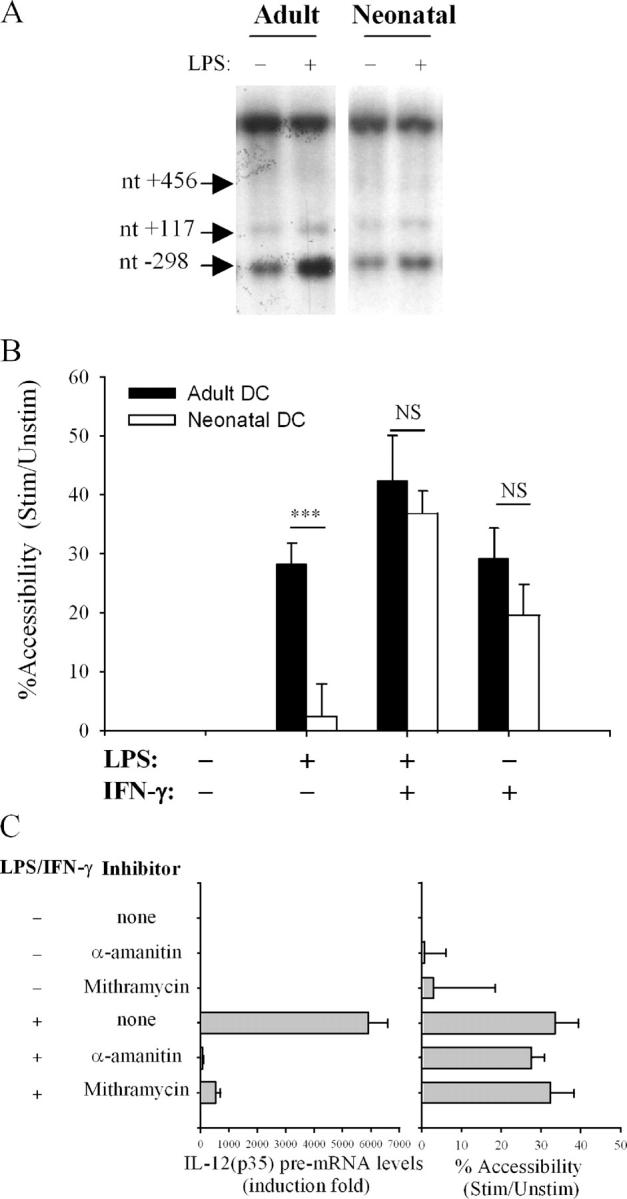
To gain insight into the inability of newborns to mount efficient Th1 responses, we analyzed the molecular basis of defective IL-12(p35) expression in human neonatal monocyte-derived dendritic cells (DCs). Determination of IL-12(p35) pre-mRNA levels by real-time RT-PCR revealed that transcriptional activation of the gene in lipopolysaccharide-stimulated neonatal DCs was strongly impaired compared with adult DCs. We next showed that p50/p65 and p65/p65 dimers interact with kB#1 site, a critical cis-acting element of the IL-12(p35) promoter. We found that LPS-induced p65 activation was similar in adult and newborn DCs. Likewise, in vitro binding activity to the Sp1#1 site, previously shown to be critical for IL-12(p35) gene activation, did not differ in adults and newborns. Since the accessibility to this Sp1#1 site was found to depend on nucleosome remodeling, we used a chromatin accessibility assay to compare remodeling of the relevant nucleosome (nuc-2) in adult and neonatal DCs. We observed that nuc-2 remodeling in neonatal DCs was profoundly impaired in response to lipopolysaccharide. Both nuc-2 remodeling and IL-12(p35) gene transcription were restored upon addition of recombinant interferon-gamma. We conclude that IL-12(p35) transcriptional repression in neonatal DCs takes place at the chromatin level.
Figures
References
-
- Wegmann, T.G., H. Lin, L. Guilbert, and T.R. Mosmann. 1993. Bidirectional cytokine interactions in the maternal-fetal relationship: is successful pregnancy a TH2 phenomenon? Immunol. Today. 14:353–356. - PubMed
-
- Siegrist, C.A. 2000. Vaccination in the neonatal period and early infancy. Int. Rev. Immunol. 19:195–219. - PubMed
-
- Holt, P.G., and C. Macaubas. 1997. Development of long-term tolerance versus sensitisation to environmental allergens during the perinatal period. Curr. Opin. Immunol. 9:782–787. - PubMed
-
- White, G.P., P.M. Watt, B.J. Holt, and P.G. Holt. 2002. Differential patterns of methylation of the IFN-gamma promoter at CpG and non-CpG sites underlie differences in IFN-gamma gene expression between human neonatal and adult CD45RO- T cells. J. Immunol. 168:2820–2827. - PubMed
-
- Hunt, D.W., H.I. Huppertz, H.J. Jiang, and R.E. Petty. 1994. Studies of human cord blood dendritic cells: evidence for functional immaturity. Blood. 84:4333–4343. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
