

Respiratory complex III is required to maintain complex I in mammalian mitochondria
- PMID: 15053874
- PMCID: PMC3164363
- DOI: 10.1016/s1097-2765(04)00124-8
Respiratory complex III is required to maintain complex I in mammalian mitochondria
Abstract
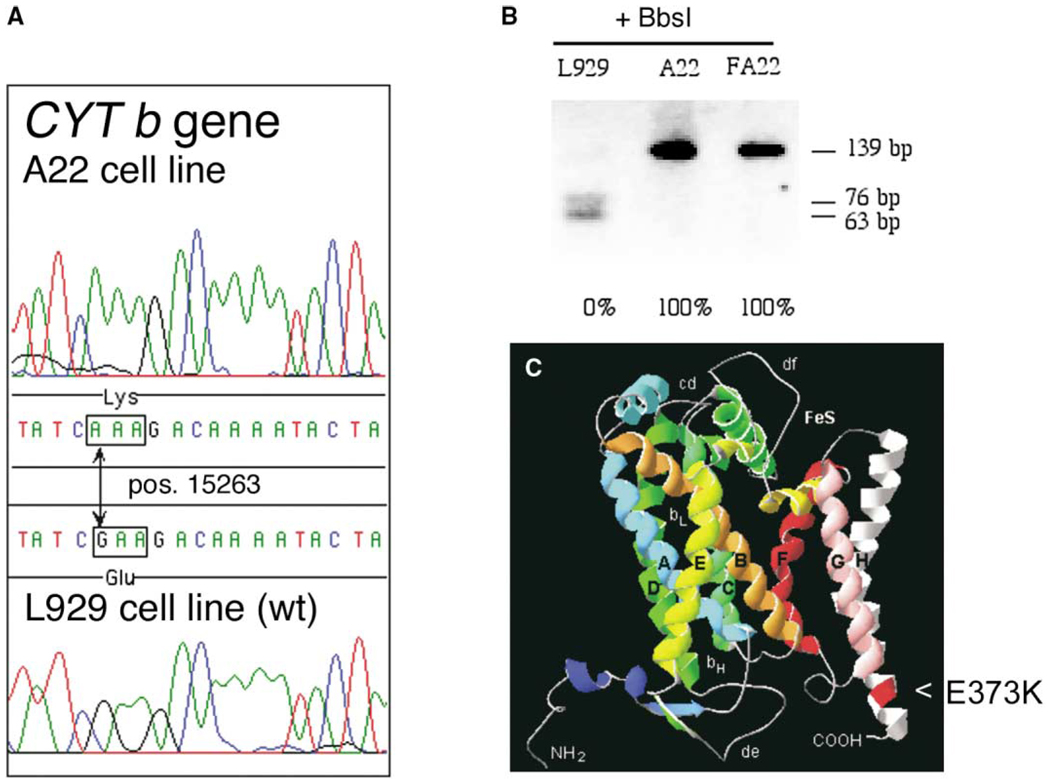
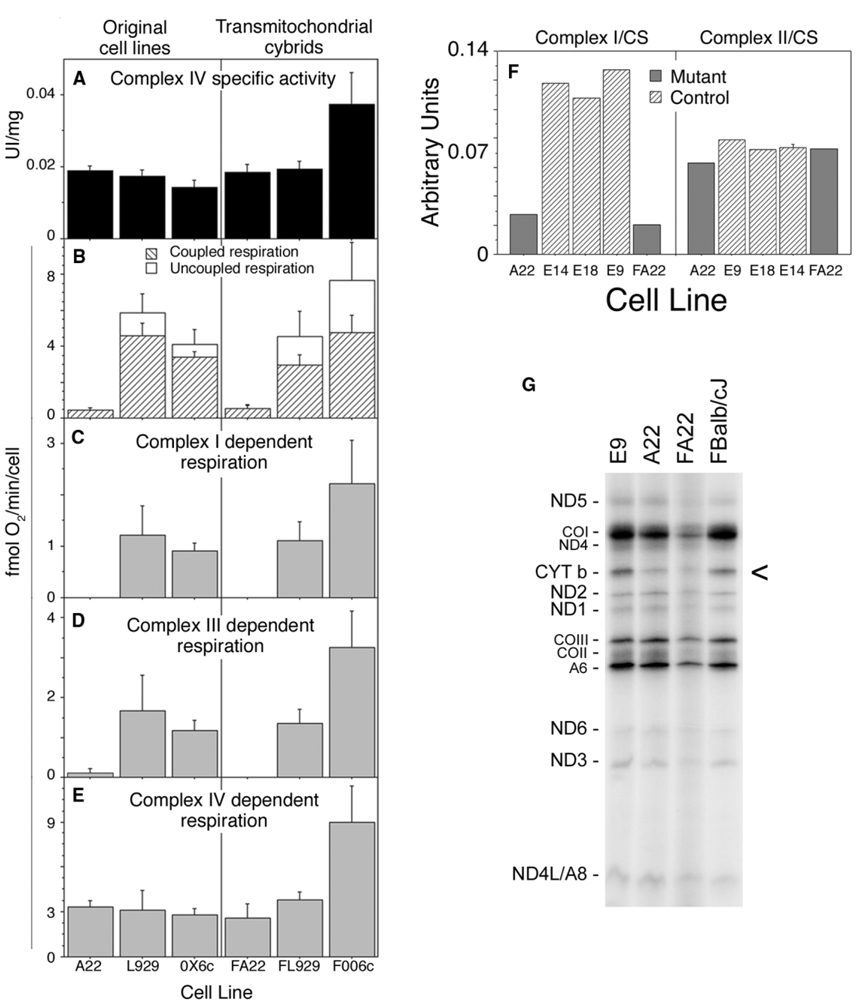
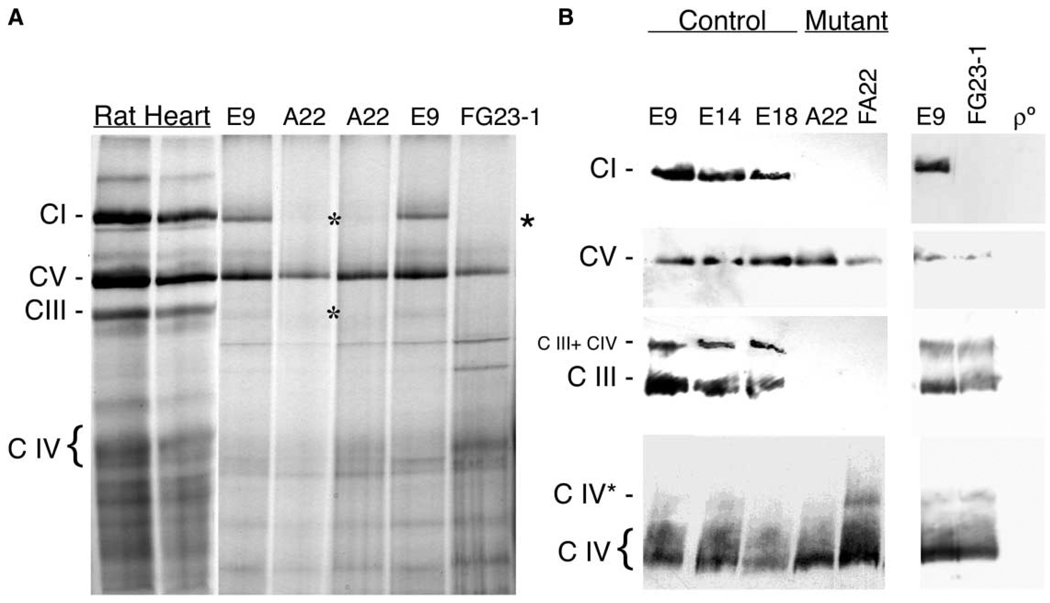
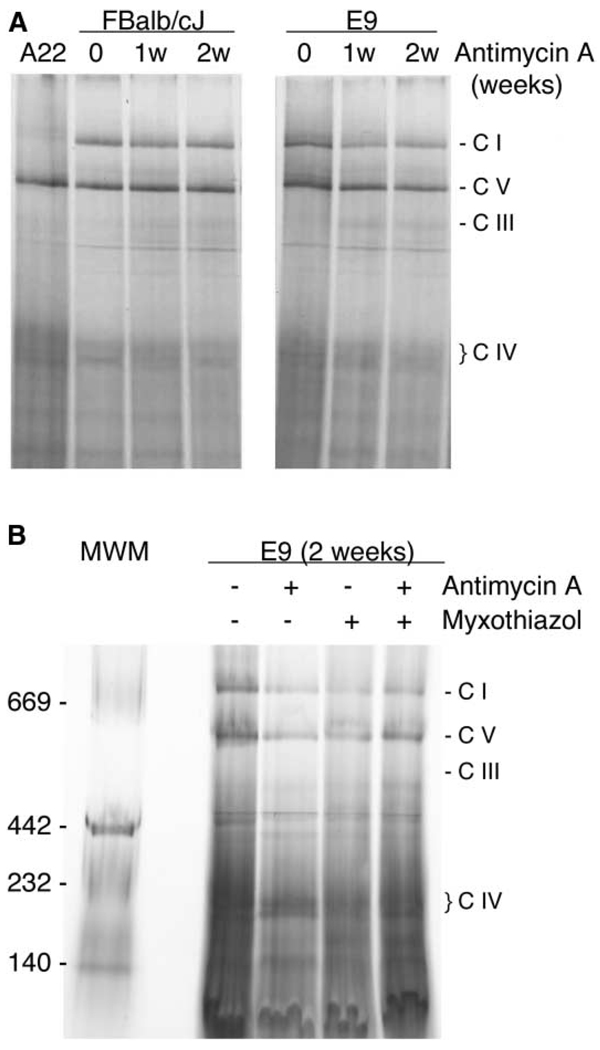
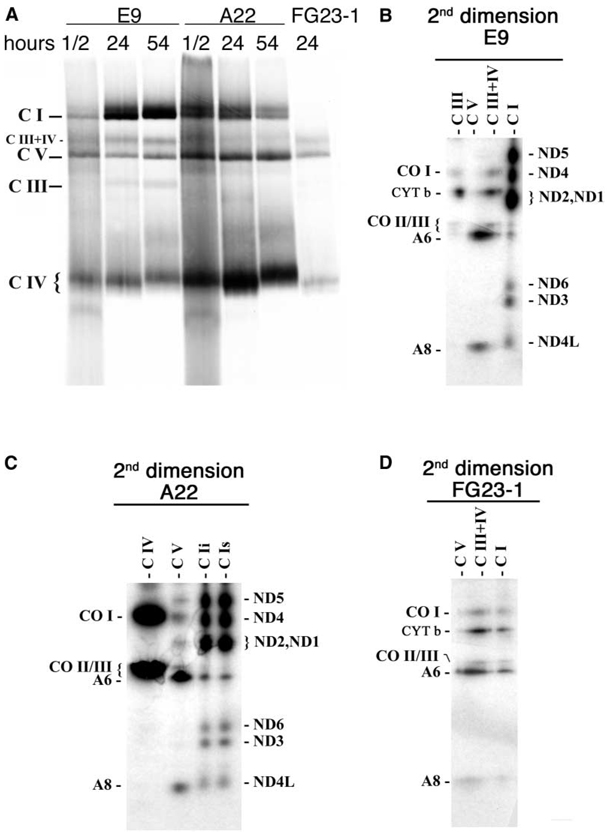
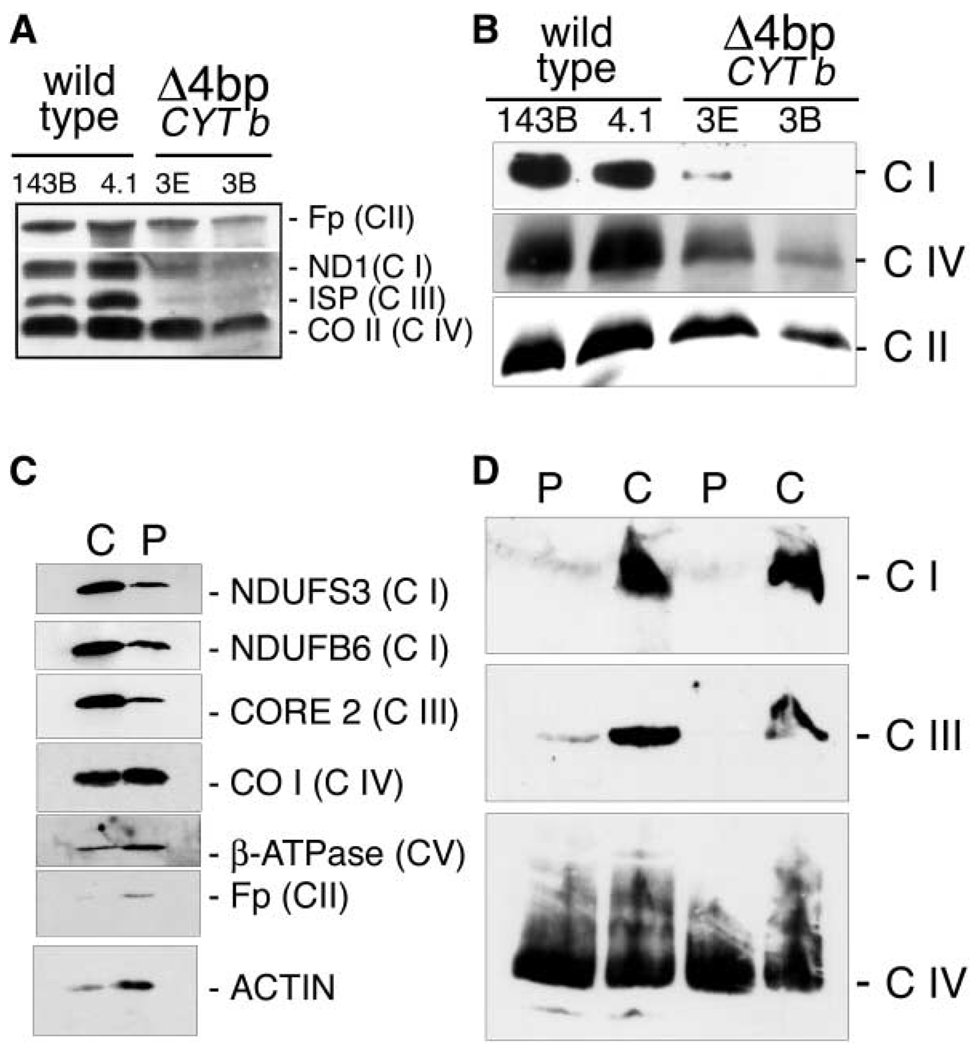
A puzzling observation in patients with oxidative phosphorylation (OXPHOS) deficiencies is the presence of combined enzyme complex defects associated with a genetic alteration in only one protein-coding gene. In particular, mutations in the mtDNA encoded cytochrome b gene are associated either with combined complex I+III deficiency or with only complex III deficiency. We have reproduced the combined complex I+III defect in mouse and human cultured cell models harboring cytochrome b mutations. In both, complex III assembly is impeded and causes a severe reduction in the amount of complex I, not observed when complex III activity was pharmacologically inhibited. Metabolic labeling in mouse cells revealed that complex I was assembled, although its stability was severely hampered. Conversely, complex III stability was not influenced by the absence of complex I. This structural dependence among complexes I and III was confirmed in a muscle biopsy of a patient harboring a nonsense cytochrome b mutation.
Figures
References
-
- Acin-Perez R, Bayona-Bafaluy MP, Bueno M, Machicado C, Fernandez-Silva P, Perez-Martos A, Montoya J, Lopez-Perez MJ, Sancho J, Enriquez JA. An intragenic suppressor in the cytochrome c oxidase I gene of mouse mitochondrial DNA. Hum. Mol. Genet. 2003;12:329–339. - PubMed
-
- Andreu AL, Bruno C, Shanske S, Shtilbans A, Hirano M, Krishna S, Hayward L, Systrom DS, Brown RH, Jr, Di-Mauro S. Missense mutation in the mtDNA cytochrome b gene in a patient with myopathy. Neurology. 1998;51:1444–1447. - PubMed
-
- Andreu AL, Hanna MG, Reichmann H, Bruno C, Penn AS, Tanji K, Pallotti F, Iwata S, Bonilla E, Lach B, et al. Exercise intolerance due to mutations in the cytochrome b gene of mitochondrial DNA. N. Engl. J. Med. 1999;341:1037–1044. - PubMed
-
- Antonicka H, Ogilvie I, Taivassalo T, Anitori RP, Haller RG, Vissing J, Kennaway NG, Shoubridge EA. Identification and characterization of a common set of complex I assembly intermediates in mitochondria from patients with complex I deficiency. J. Biol. Chem. 2003;278:43081–43088. - PubMed
-
- Appleby RD, Porteous WK, Hughes G, James AM, Shannon D, Wei YH, Murphy MP. Quantitation and origin of the mitochondrial membrane potential in human cells lacking mitochondrial DNA. Eur. J. Biochem. 1999;262:108–116. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
