

doi: 10.1073/pnas.0307247101.
Epub 2004 Mar 31.
Myosin V processivity: multiple kinetic pathways for head-to-head coordination
Affiliations
- PMID: 15056760
- PMCID: PMC397419
- DOI: 10.1073/pnas.0307247101
Item in Clipboard
Myosin V processivity: multiple kinetic pathways for head-to-head coordination
Proc Natl Acad Sci U S A.
.
Abstract
Myosin V, a double-headed molecular motor, transports organelles within cells by walking processively along actin, a process that requires coordination between the heads. To understand the mechanism underlying this coordination, processive runs of single myosin V molecules were perturbed by varying nucleotide content. Contrary to current views, our results show that the two heads of a myosin V molecule communicate, not through any one mechanism but through an elaborate system of cooperative mechanisms involving multiple kinetic pathways. These mechanisms introduce redundancy and safeguards that ensure robust processivity under differing physiologic demands.
Figures
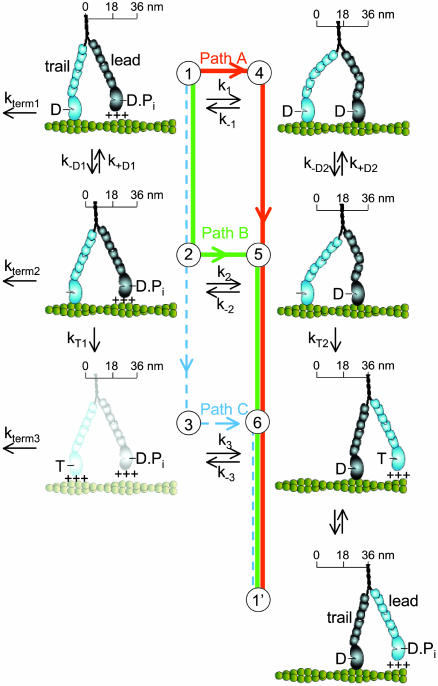
A simple six-state model showing the multiple pathways that the two heads of a myosin V molecule might follow during their individual ATPase cycles as they process along an actin filament (green helical structure). The scheme represents only half of a cycle for a given head (the second half would have the heads swapping roles as lead and trail heads, states 6 → 1′) and is arranged such that transitions from top to bottom are the trail head's (light blue) biochemical transitions (ADP release and ATP binding), with transitions from left to right being the lead head's (dark blue) working step. The working step involves a relatively discrete rotation of the light-chain domain of myosin (six small ovals) relative to its catalytic domain (large oval) and is closely associated with Pi release and strong binding to actin (illustrated as the catalytic domain making contact with the actin filament). Weak binding is indicated by the +++ signs and is assumed to be a rapid, nonspecific attachment and detachment of the myosin head to actin. The working step can be partitioned between movement (illustrated as translocation along a scale) and the generation of intramolecular strain (illustrated as the bending of the myosin light-chain domain). An additional light-chain domain rotation associated with ADP release (26, 27) results in additional movement (state 1 → 2) and an energetically favorable relaxation of any intramolecular strain (state 4 → 5). The complete relaxation of intramolecular strain after ATP binding may result in additional movement (state 5 → 6). Finally, termination of a processive run, at a rate of kterm 1, is assumed to occur from a state at which only one ADP-bound head is attached to actin. Although myosin V might terminate from state 6, kinetic studies (13) indicate that the lifetime of this state (<5 msec) is short relative to the lifetime of state 1, making the probability of detachment from this state relatively low.
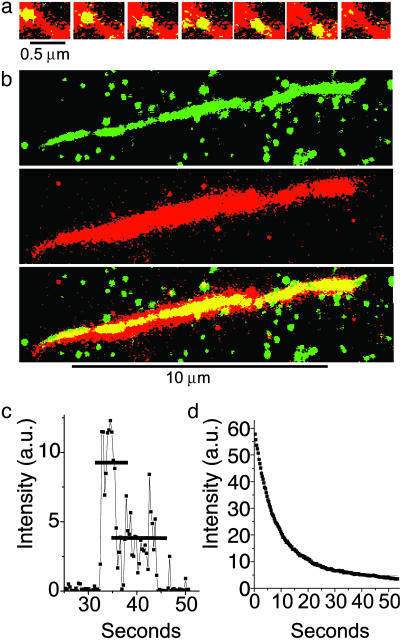
(a) A series of images showing a single YFP-HMMM5 molecule moving along an actin filament. The Alexa 660/biotin-actin filament (red) was first imaged in epifluorescence. At the same location, a series of YFP-HMMM5 images (green) were subsequently obtained at 0.3-sec intervals in TIRF mode. The stacked red and green images show regions of overlap (colocalization) as yellow, thus highlighting the position and movement of YFP-HMMM5 on actin. YFP-HMMM5 movement was always unidirectional, suggesting that movement occurred on single actin filaments. (b) YFP-HMMM5 molecules travel the full length of the actin filament. A 120-frame average (≈36 sec) of ≈40 YFP-HMMM5 processive runs (Top, green, imaged with TIRF) along an actin filament (Middle, red, imaged with epifluorescence). The stacked images (Bottom), with regions of overlap shown in yellow, demonstrate our observation that as the YFP-HMMM5 travels along the actin filament, it had access to the entire length of actin, suggesting that few inaccessible regions exist. The green spots in the Top and Bottom images are YFP-HMMM5 molecules that are bound nonspecifically to the surface at which actin was not present. (c) Time course of fluorescence intensity [arbitrary units (a.u.)] showing photobleaching of a single YFP-HMMM5 molecule. Most (>95%) YFP-HMMM5 molecules photobleached with either one or two steps, the latter (shown here) corresponding to the photobleaching of two YFP molecules, one bound to the C terminus of each of the two heavy chains of HMMM5. The absence of photobleaching events that occur in more than two steps indicates that the fluorescent images originated from a single myosin V molecule and not from an aggregate. (d) Time course of fluorescence intensity (arbitrary units) showing photobleaching of many YFP-HMMM5 molecules. A single exponential fitofthe fluorescence decay (red line) of many YFP-HMMM5 molecules bound to actin filaments in rigor gave a photobleaching rate of ≈0.1 sec–1, which is significantly slower than the range of inverse run times reported in this article (0.17–0.7 sec–1), indicating that photobleaching kinetics do not influence our run-time estimates significantly. Indeed, including the photobleaching rates in our analysis did not change our conclusions.
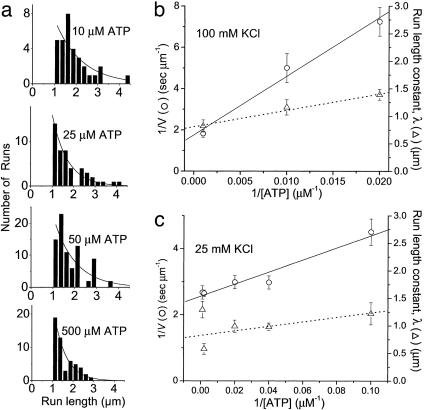
Velocities and run lengths versus [ATP]. (a) Run lengths, L, observed at four different ATP concentrations (10, 25, 50, and 500 μm) are plotted as histograms. Solid lines are Ae–L/λ, where λ is the run-length constant. Inverse velocity (1/V, ○) and run lengths constant (▵) are plotted versus 1/[ATP] for a single YFP-HMMM5 at 100 mM KCl (b) and 25 mM KCl (c). The solid and dashed lines are linear regressions of the velocity and run-length data, respectively.
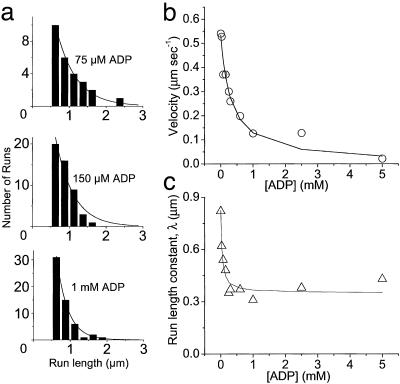
Velocities and run lengths versus [ADP] at 1 mM ATP and 100 mM KCl. (a) Run lengths, L, observed at three different ADP concentrations (75, 150, and 1,000 μM) are plotted as histograms. Solid lines are Ae–L/λ, where λ is the run-length constant. (b) Velocities are plotted versus [ADP] for a single YFP-HMMM5. The solid line is the best fit of Eq. 1 (substituting in Eq. 2) to the velocity data. (c) Run-length constants are plotted versus [ADP]. The solid line is the best fitofEq. 4 to the run-length data.
References
-
- Mehta, A. D., Rock, R. S., Rief, M., Spudich, J. A., Mooseker, M. S. & Cheney, R. E. (1999) Nature 400, 590–593. - PubMed
-
- Sakamoto, T., Amitani, I., Yokota, E. & Ando, T. (2000) Biochem. Biophys. Res. Commun. 272, 586–590. - PubMed
-
- Pastural, E., Barrat, F. J., Dufourcq-Lagelouse, R., Certain, S., Sanal, O., Jabado, N., Seger, R., Griscelli, C., Fischer, A. & de Saint, B. G. (1997) Nat. Genet. 16, 289–292. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
