

Hepatic expansion of a virus-specific regulatory CD8(+) T cell population in chronic hepatitis C virus infection
- PMID: 15057302
- PMCID: PMC379326
- DOI: 10.1172/JCI20515
Hepatic expansion of a virus-specific regulatory CD8(+) T cell population in chronic hepatitis C virus infection
Abstract
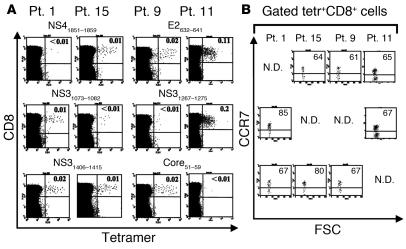
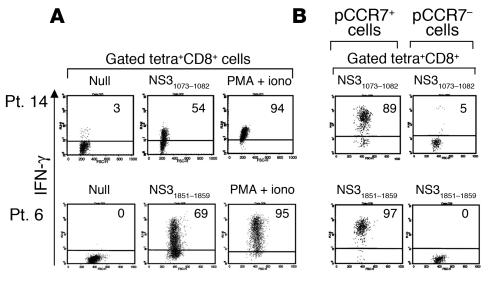
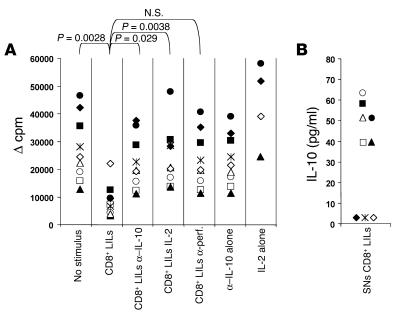
Regulatory T (T(R)) cells consist of phenotypically and functionally distinct CD4(+) and CD8(+) T cell subsets engaged both in maintaining self-tolerance and in preventing anti-non-self effector responses (microbial, tumor, transplant, and so on) that may be harmful to the host. Here we propose that the proinflammatory function of virus-specific memory effector CCR7(-)CD8(+) T cells, which are massively recruited in the liver, are inefficient (in terms of IFN-gamma production) in patients with chronic hepatitis C virus (HCV) infection because of the concomitant presence of virus-specific CCR7(-)CD8(+) T(R) cells producing considerable amounts of IL-10. These CD8(+) T(R) cells are antigen specific, as they can be stimulated by HCV epitopes and suppress T cell responses that are in turn restored by the addition of neutralizing anti-IL-10. This study provides for the first time to our knowledge direct evidence of the existence of virus-specific CD8(+) T(R) cells that infiltrate the livers of patients with chronic HCV infection, identifies IL-10 as a soluble inhibitory factor mediating suppression, and suggests that these cells play a pivotal role in controlling hepatic effector CD8(+) T cell responses.
Figures





References
-
- Lauer GM, Walker BD. Hepatitis C virus infection. N. Engl. J. Med. 2001;345:41–52. - PubMed
-
- Cooper S, et al. Analysis of a successful immune response against hepatitis C virus. Immunity. 1999;10:439–449. - PubMed
-
- Prezzi C, et al. Virus-specific CD8+ T cells with type 1 or type 2 cytokine profile are related to different disease activity in chronic hepatitis C virus infection. Eur. J. Immunol. 2001;31:894–906. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
