

Transcriptional organization and posttranscriptional regulation of the Bacillus subtilis branched-chain amino acid biosynthesis genes
- PMID: 15060025
- PMCID: PMC412147
- DOI: 10.1128/JB.186.8.2240-2252.2004
Transcriptional organization and posttranscriptional regulation of the Bacillus subtilis branched-chain amino acid biosynthesis genes
Abstract
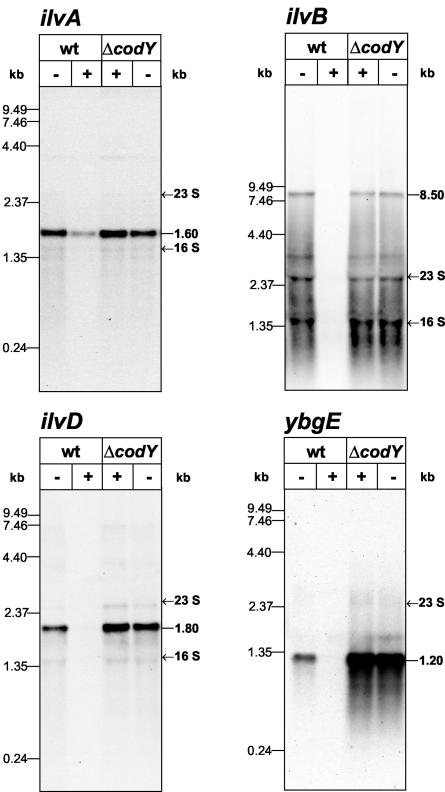
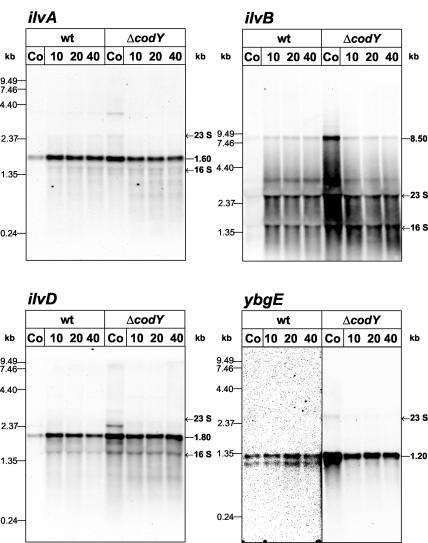
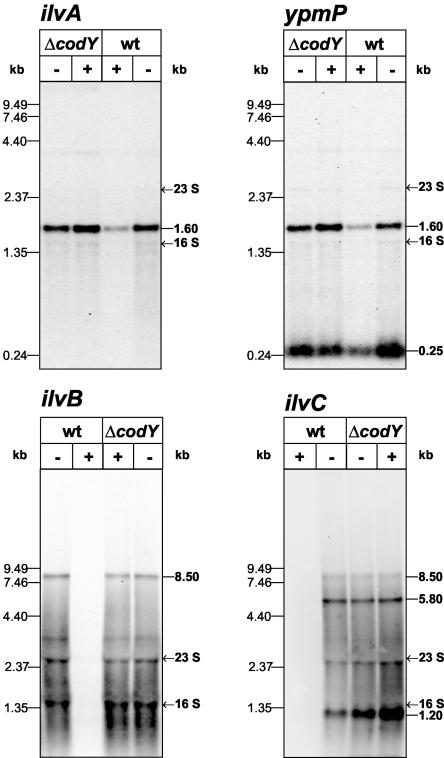
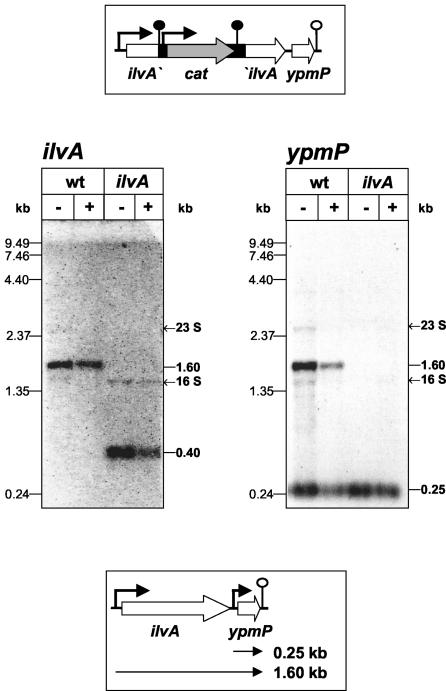
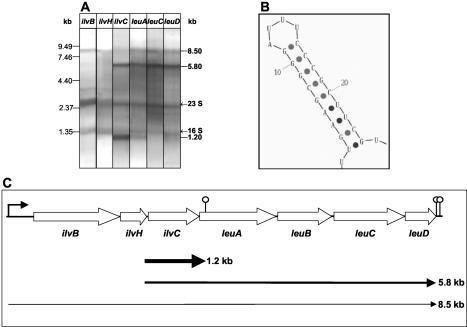
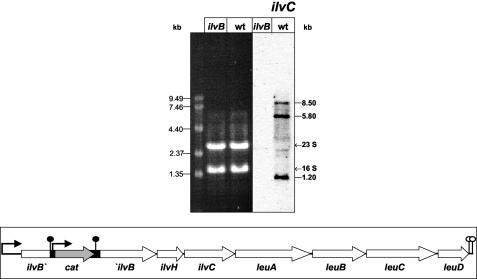
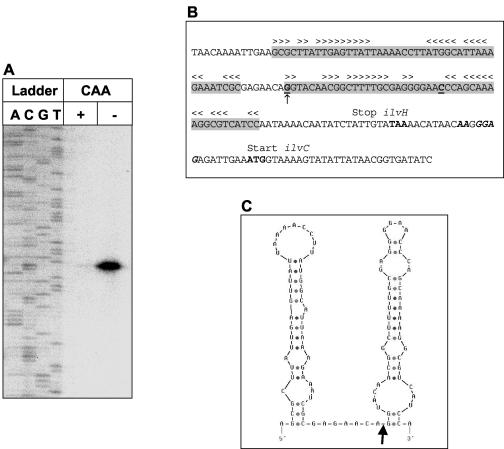
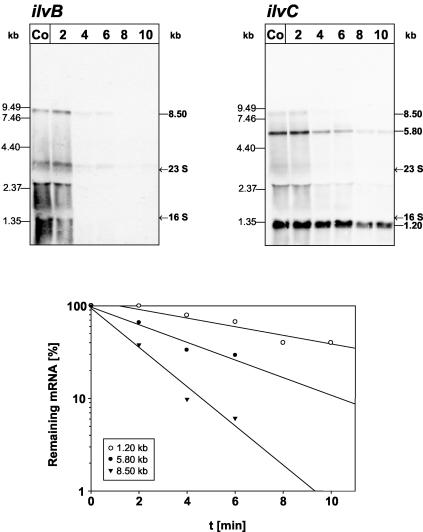
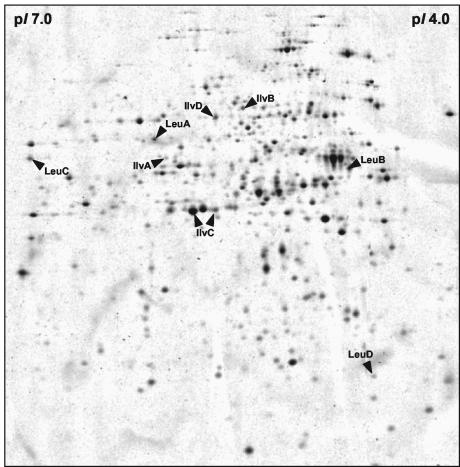
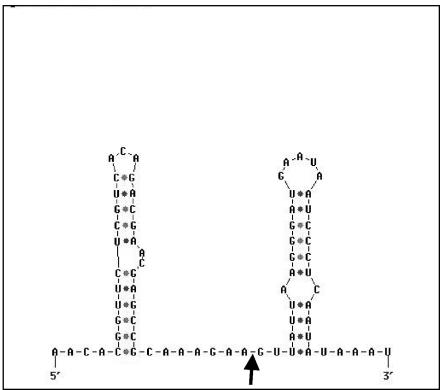
In Bacillus subtilis, the genes of the branched-chain amino acids biosynthetic pathway are organized in three genetic loci: the ilvBHC-leuABCD (ilv-leu) operon, ilvA, and ilvD. These genes, as well as ybgE, encoding a branched-chain amino acid aminotransferase, were recently demonstrated to represent direct targets of the global transcriptional regulator CodY. In the present study, the transcriptional organization and posttranscriptional regulation of these genes were analyzed. Whereas ybgE and ilvD are transcribed monocistronically, the ilvA gene forms a bicistronic operon with the downstream located ypmP gene, encoding a protein of unknown function. The ypmP gene is also directly preceded by a promoter sharing the regulatory pattern of the ilvA promoter. The ilv-leu operon revealed complex posttranscriptional regulation: three mRNA species of 8.5, 5.8, and 1.2 kb were detected. Among them, the 8.5-kb full-length primary transcript exhibits the shortest half-life (1.2 min). Endoribonucleolytic cleavage of this transcript generates the 5.8-kb mRNA, which lacks the coding sequences of the first two genes of the operon and is predicted to carry a stem-loop structure at its 5' end. This processing product has a significantly longer half-life (3 min) than the full-length precursor. The most stable transcript (half-life, 7.6 min) is the 1.2-kb mRNA generated by the processing event and exonucleolytic degradation of the large transcripts or partial transcriptional termination. This mRNA, which encompasses exclusively the ilvC coding sequence, is predicted to carry a further stable stem-loop structure at its 3' end. The very different steady-state amounts of mRNA resulting from their different stabilities are also reflected at the protein level: proteome studies revealed that the cellular amount of IlvC protein is 10-fold greater than that of the other proteins encoded by the ilv-leu operon. Therefore, differential segmental stability resulting from mRNA processing ensures the fine-tuning of the expression of the individual genes of the operon.
Figures
References
-
- Belasco, J. G., J. T. Beatty, C. W. Adams, A. von Gabain, and S. N. Cohen. 1985. Differential expression of photosynthesis genes in R. capsulata results from segmental differences in stability within the polycistronic rxcA transcript. Cell 40:171-181. - PubMed
-
- Biou, V., R. Dumas, C. Cohen-Addad, R. Douce, D. Job, and E. Pebay-Peyroula. 1997. The crystal structure of plant acetohydroxy acid isomeroreductase complexed with NADPH, two magnesium ions and a herbicidal transition state analog determined at 1.65 A resolution. EMBO J. 16:3405-3415. - PMC - PubMed
-
- Büttner, K., J. Bernhardt, C. Scharf, R. Schmid, U. Mäder, C. Eymann, H. Antelmann, A. Völker, U. Völker, and M. Hecker. 2001. A comprehensive two-dimensional map of cytosolic proteins of Bacillus subtilis. Electrophoresis 22:2908-2935. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
