

Genetic analysis of the Salmonella enterica type III secretion-associated ATPase InvC defines discrete functional domains
- PMID: 15060043
- PMCID: PMC412161
- DOI: 10.1128/JB.186.8.2402-2412.2004
Genetic analysis of the Salmonella enterica type III secretion-associated ATPase InvC defines discrete functional domains
Abstract
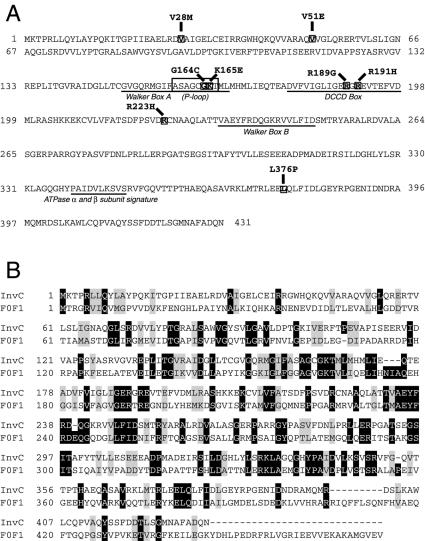
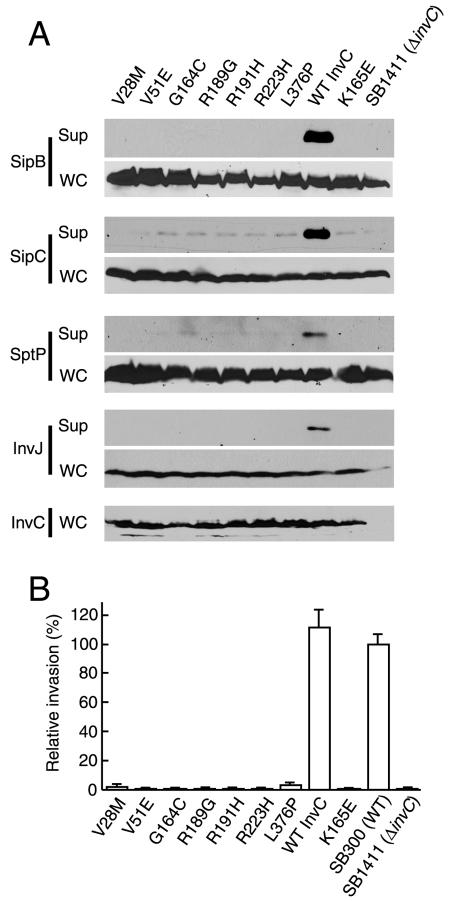
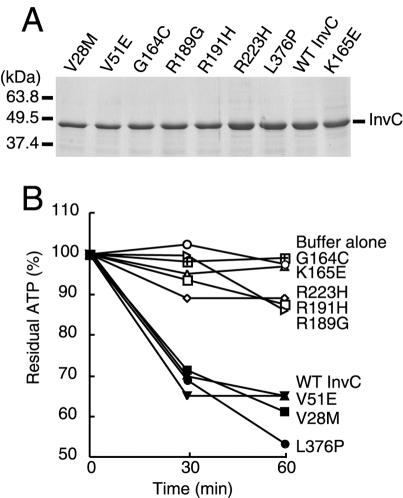
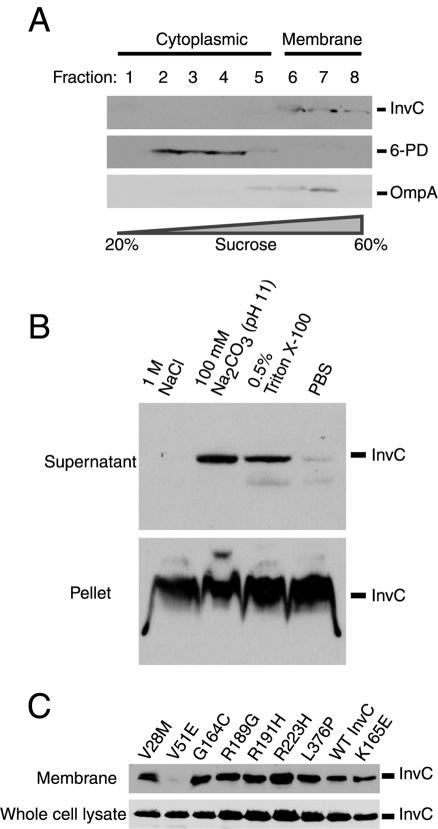
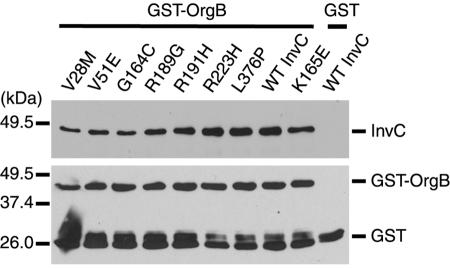
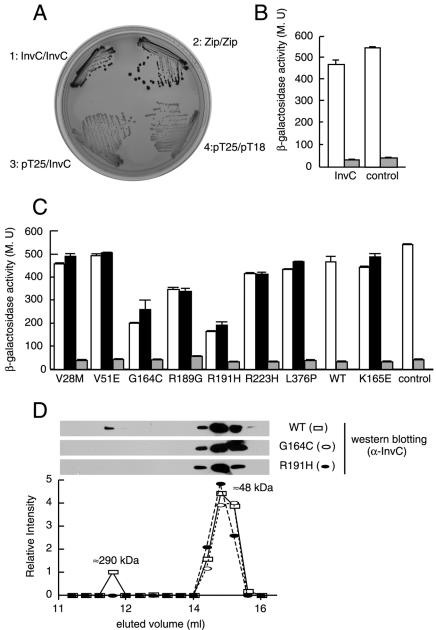
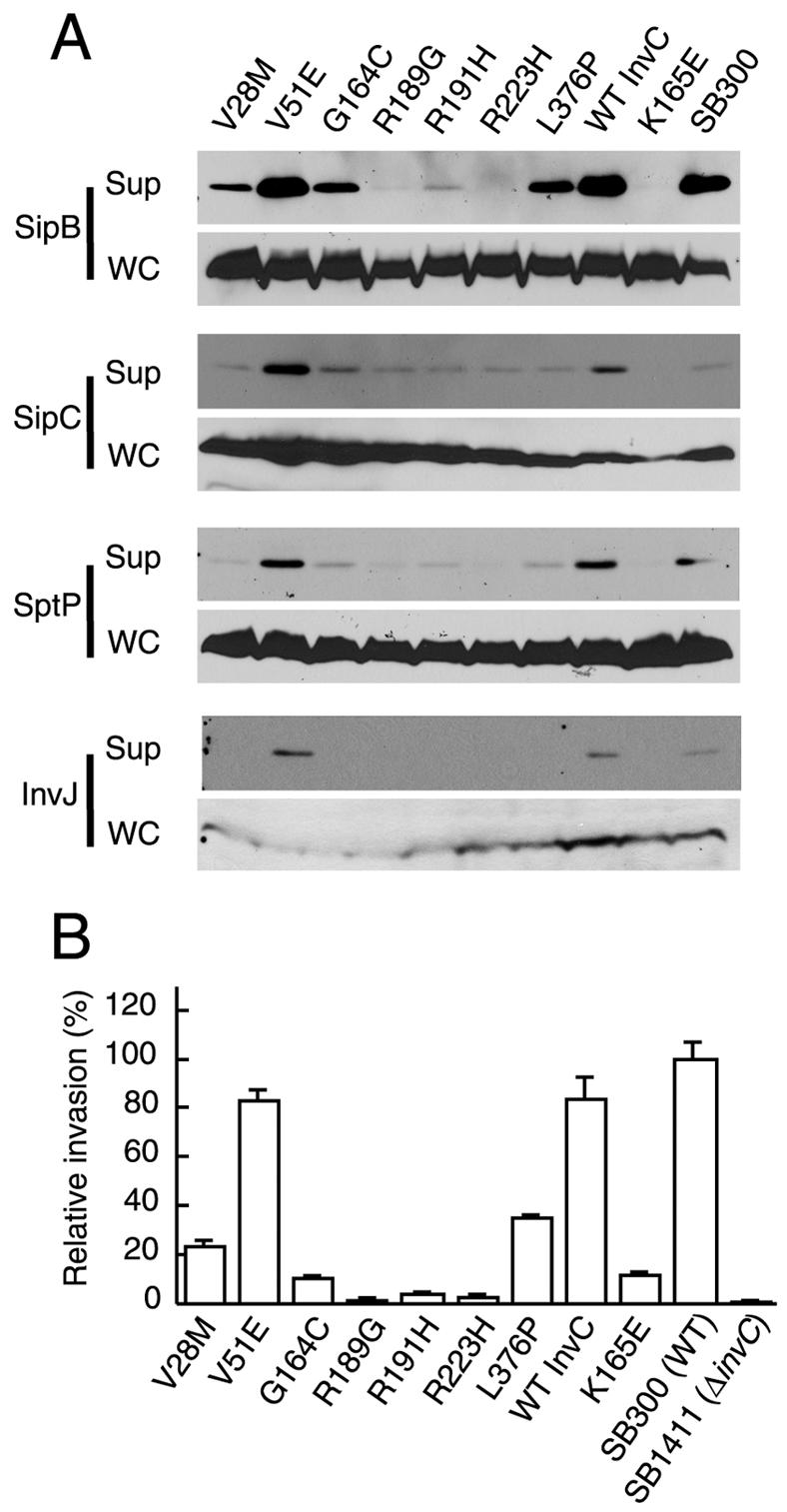
An essential component of all type III secretion systems is a highly conserved ATPase that shares significant amino acid sequence similarity to the beta subunit of the F(0)F(1) ATPases and is thought to provide the energy for the secretion process. We have performed a genetic and functional analysis of InvC, the ATPase associated with the Salmonella enterica type III secretion system encoded within its pathogenicity island 1. Through a mutagenesis analysis, we have identified amino acid residues that are essential for specific activities of InvC, such as nucleotide hydrolysis and membrane binding. This has allowed us to define discrete domains of InvC that are specifically associated with different essential activities of this protein.
Figures

References
-
- Boyer, P. 1997. The ATP synthase—a splendid molecular machine. Annu. Rev. Biochem. 66:717-749. - PubMed
-
- Brusilow, W. S. A. 1993. Assembly of the Escherichia coli F1F0 ATPase, a large multimeric membrane-bound enzyme. Mol. Microbiol. 9:419-424. - PubMed
-
- Collazo, C. M., M. K. Zierler, and J. E. Galán. 1995. Functional analysis of the Salmonella typhimurium invasion genes invI and invJ and identification of a target of the protein secretion apparatus encoded in the inv locus. Mol. Microbiol. 15:25-38. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
