

In vivo dynamics of Swi6 in yeast: evidence for a stochastic model of heterochromatin
- PMID: 15060140
- PMCID: PMC381678
- DOI: 10.1128/MCB.24.8.3157-3167.2004
In vivo dynamics of Swi6 in yeast: evidence for a stochastic model of heterochromatin
Abstract
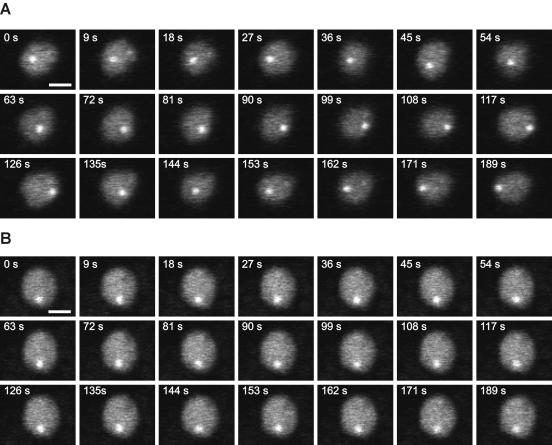
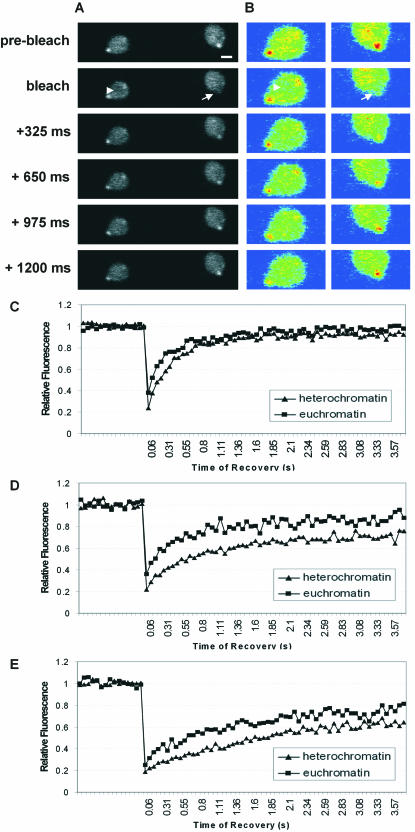
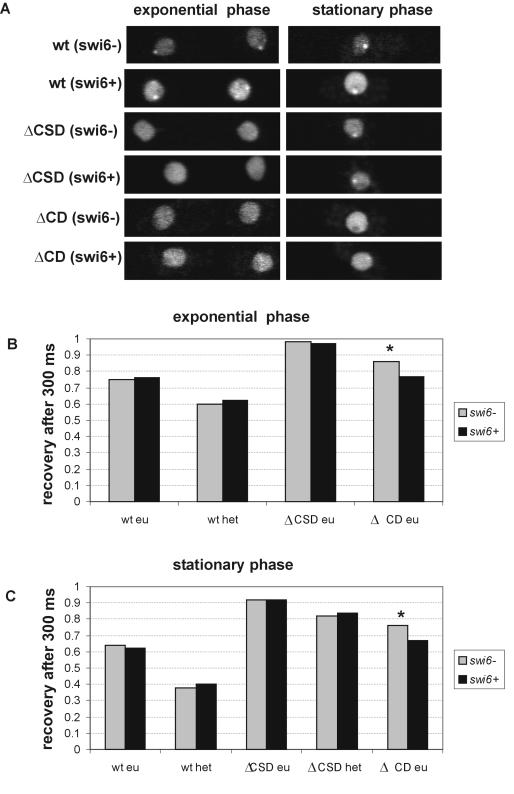
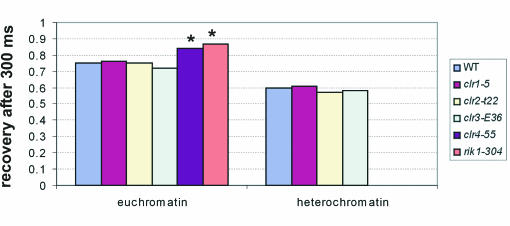
The mechanism for transcriptional silencing of pericentric heterochromatin is conserved from fission yeast to mammals. Silenced genome regions are marked by epigenetic methylation of histone H3, which serves as a binding site for structural heterochromatin proteins. In the fission yeast Schizosaccharomyces pombe, the major structural heterochromatin protein is Swi6. To gain insight into Swi6 function in vivo, we have studied its dynamics in the nucleus of living yeast. We demonstrate that, in contrast to mammalian cells, yeast heterochromatin domains undergo rapid, large-scale motions within the nucleus. Similar to the situation in mammalian cells, Swi6 does not permanently associate with these chromatin domains but binds only transiently to euchromatin and heterochromatin. Swi6 binding dynamics are dependent on growth status and on the silencing factors Clr4 and Rik1, but not Clr1, Clr2, or Clr3. By comparing the kinetics of mutant Swi6 proteins in swi6(-) and swi6(+) strains, we demonstrate that homotypic protein-protein interactions via the chromoshadow domain stabilize Swi6 binding to chromatin in vivo. Kinetic modeling allowed quantitative estimation of residence times and indicated the existence of at least two kinetically distinct populations of Swi6 in heterochromatin. The observed dynamics of Swi6 binding are consistent with a stochastic model of heterochromatin and indicate evolutionary conservation of heterochromatin protein binding properties from mammals to yeast.
Figures
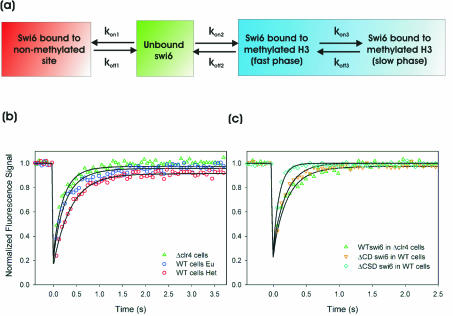
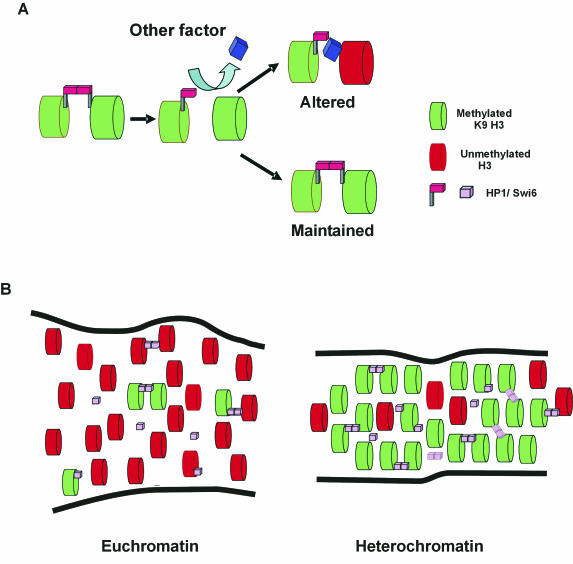
References
-
- Ahmad, K., and S. Henikoff. 2001. Modulation of a transcription factor counteracts heterochromatic gene silencing in Drosophila. Cell 104:839-847. - PubMed
-
- Allshire, R. C., E. R. Nimmo, K. Ekwall, J. P. Javerzat, and G. Cranston. 1995. Mutations derepressing silent centromeric domains in fission yeast disrupt chromosome segregation. Genes Dev. 9:218-233. - PubMed
-
- Aparicio, O. M., and D. E. Gottschling. 1994. Overcoming telomeric silencing: a trans-activator competes to establish gene expression in a cell cycle-dependent way. Genes Dev. 8:1133-1146. - PubMed
-
- Bannister, A. J., R. Schneider, and T. Kouzarides. 2002. Histone methylation: dynamic or static? Cell 109:801-806. - PubMed
-
- Bannister, A. J., P. Zegerman, J. F. Partridge, E. A. Miska, J. O. Thomas, R. C. Allshire, and T. Kouzarides. 2001. Selective recognition of methylated lysine 9 on histone H3 by the HP1 chromo domain. Nature 410:120-124. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources