

Presynaptic inhibition of spontaneous acetylcholine release induced by adenosine at the mouse neuromuscular junction
- PMID: 15066904
- PMCID: PMC1574912
- DOI: 10.1038/sj.bjp.0705656
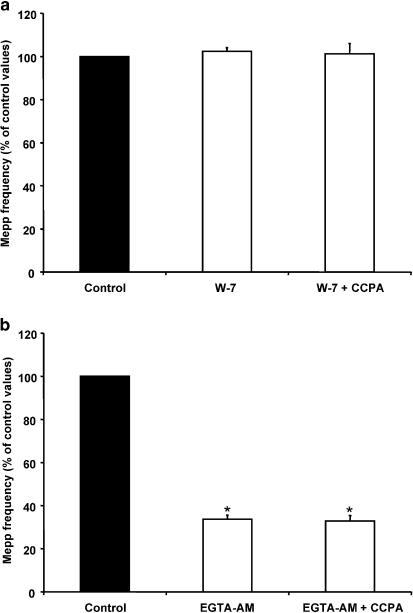
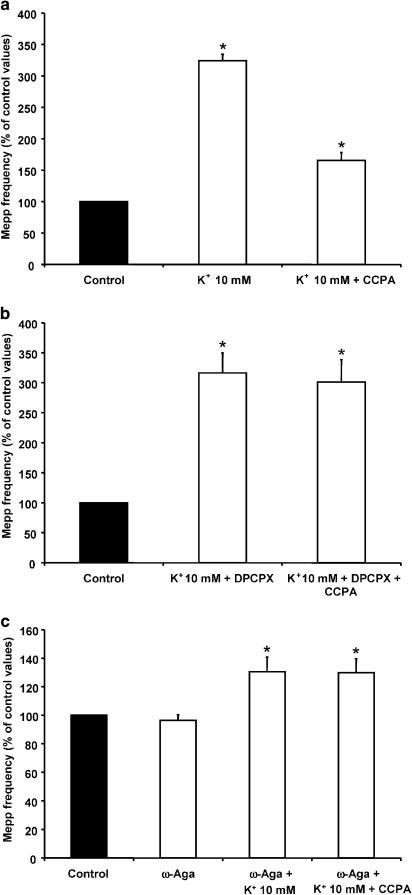
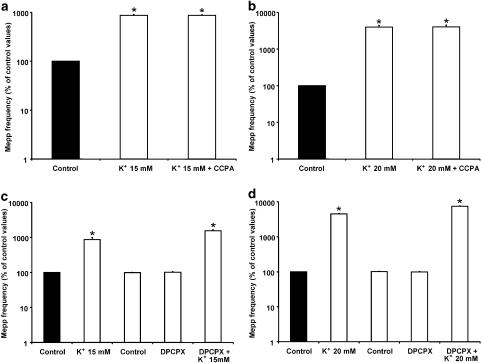
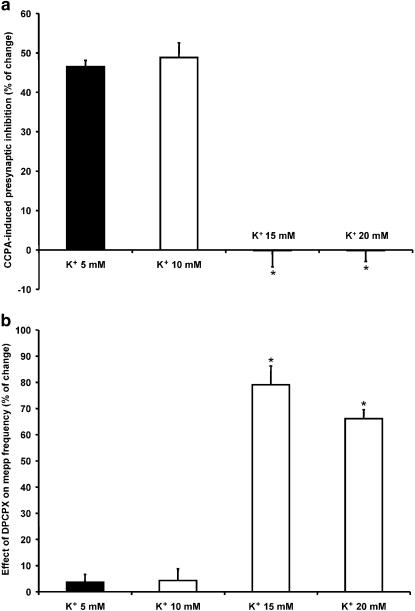
Presynaptic inhibition of spontaneous acetylcholine release induced by adenosine at the mouse neuromuscular junction
Abstract
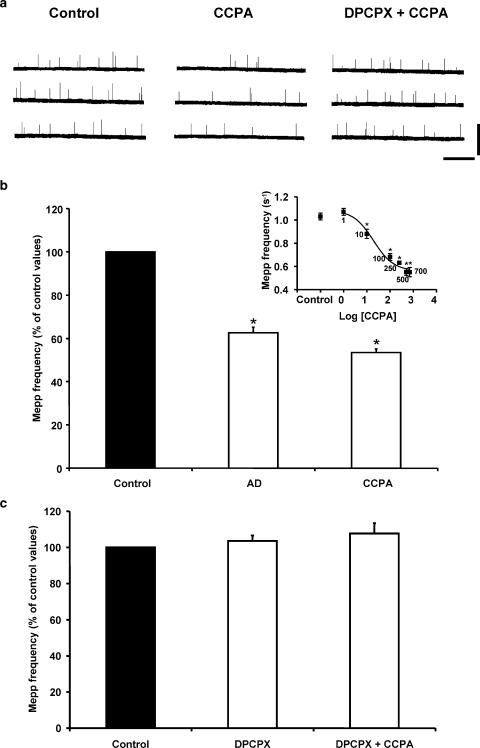
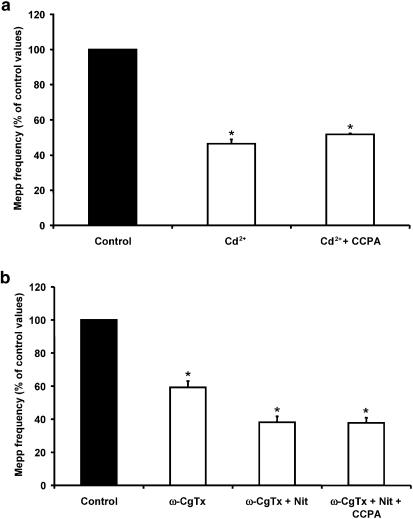
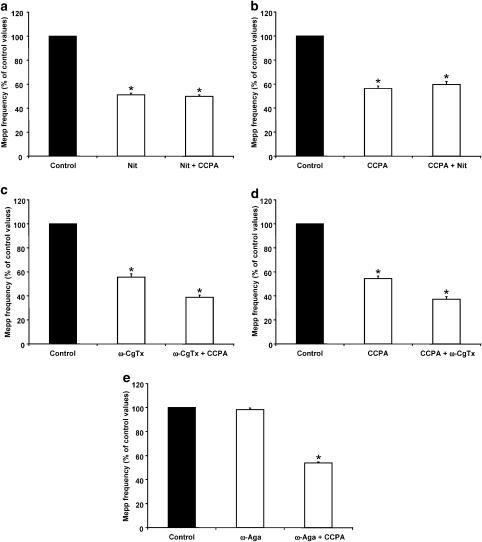
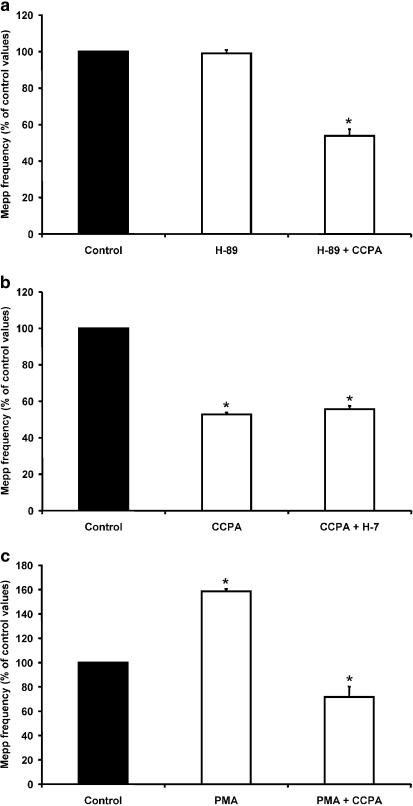
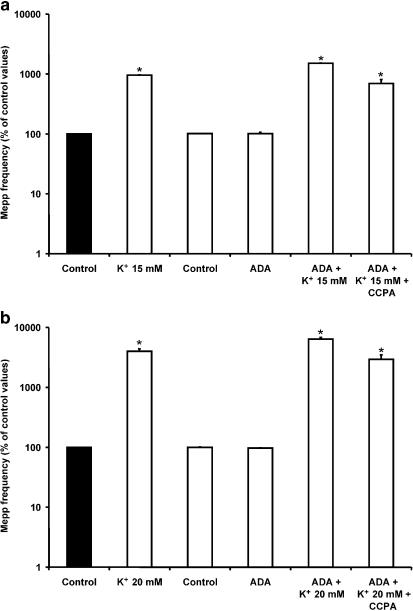
1. At the mouse neuromuscular junction, adenosine (AD) and the A(1) agonist 2-chloro-N(6)-cyclopentyl-adenosine (CCPA) induce presynaptic inhibition of spontaneous acetylcholine (ACh) release by activation of A(1) AD receptors through a mechanism that is still unknown. To evaluate whether the inhibition is mediated by modulation of the voltage-dependent calcium channels (VDCCs) associated with tonic secretion (L- and N-type VDCCs), we measured the miniature end-plate potential (mepp) frequency in mouse diaphragm muscles. 2. Blockade of VDCCs by Cd(2+) prevented the effect of the CCPA. Nitrendipine (an L-type VDCC antagonist) but not omega-conotoxin GVIA (an N-type VDCC antagonist) blocked the action of CCPA, suggesting that the decrease in spontaneous mepp frequency by CCPA is associated with an action on L-type VDCCs only. 3. As A(1) receptors are coupled to a G(i/o) protein, we investigated whether the inhibition of PKA or the activation of PKC is involved in the presynaptic inhibition mechanism. Neither N-(2[p-bromocinnamylamino]-ethyl)-5-isoquinolinesulfonamide (H-89, a PKA inhibitor), nor 1-(5-isoquinolinesulfonyl)-2-methyl-piperazine (H-7, a PKC antagonist), nor phorbol 12-myristate 13-acetate (PHA, a PKC activator) modified CCPA-induced presynaptic inhibition, suggesting that these second messenger pathways are not involved. 4. The effect of CCPA was eliminated by the calmodulin antagonist N-(6-aminohexil)-5-chloro-1-naphthalenesulfonamide hydrochloride (W-7) and by ethylene glycol-bis(beta-aminoethyl ether)-N,N,N',N'-tetraacetic acid-acetoxymethyl ester epsilon6TDelta-BM, which suggests that the action of CCPA to modulate L-type VDCCs may involve Ca(2+)-calmodulin. 5. To investigate the action of CCPA on diverse degrees of nerve terminal depolarization, we studied its effect at different external K(+) concentrations. The effect of CCPA on ACh secretion evoked by 10 mm K(+) was prevented by the P/Q-type VDCC antagonist omega-agatoxin IVA. 6. CCPA failed to inhibit the increases in mepp frequency evoked by 15 and 20 mm K(+). We demonstrated that, at high K(+) concentrations, endogenous AD occupies A1 receptors, impairing the action of CCPA, since incubation with 8-cyclopentyl-1,3-dipropylxanthine (DPCPX, an A(1) receptor antagonist) and adenosine deaminase (ADA), which degrades AD into the inactive metabolite inosine, increased mepp frequency compared with that obtained in 15 and 20 mm K(+) in the absence of the drugs. Moreover, CCPA was able to induce presynaptic inhibition in the presence of ADA. It is concluded that, at high K(+) concentrations, the activation of A(1) receptors by endogenous AD prevents excessive neurotransmitter release.
Figures
Similar articles
-
Excitatory effect of the A2A adenosine receptor agonist CGS-21680 on spontaneous and K+-evoked acetylcholine release at the mouse neuromuscular junction.Neuroscience. 2011 Jan 13;172:164-76. doi: 10.1016/j.neuroscience.2010.10.015. Epub 2010 Nov 2. Neuroscience. 2011. PMID: 20971164
-
Presynaptic inhibition of spontaneous acetylcholine release mediated by P2Y receptors at the mouse neuromuscular junction.Neuroscience. 2006 Sep 29;142(1):71-85. doi: 10.1016/j.neuroscience.2006.05.062. Epub 2006 Jul 14. Neuroscience. 2006. PMID: 16843602
-
Effect of purines on calcium-independent acetylcholine release at the mouse neuromuscular junction.Neuroscience. 2008 Jul 17;154(4):1324-36. doi: 10.1016/j.neuroscience.2008.04.056. Epub 2008 May 3. Neuroscience. 2008. PMID: 18534762
-
Presynaptic membrane receptors in acetylcholine release modulation in the neuromuscular synapse.J Neurosci Res. 2014 May;92(5):543-54. doi: 10.1002/jnr.23346. Epub 2014 Jan 27. J Neurosci Res. 2014. PMID: 24464361 Review.
-
[Molecular mechanism of exocytosis in neural and immune system].Yakugaku Zasshi. 2000 Dec;120(12):1419-27. doi: 10.1248/yakushi1947.120.12_1419. Yakugaku Zasshi. 2000. PMID: 11193390 Review. Japanese.
Cited by
-
Constitutive and evoked release of ATP in adult mouse olfactory epithelium.Open Life Sci. 2024 Jan 16;19(1):20220811. doi: 10.1515/biol-2022-0811. eCollection 2024. Open Life Sci. 2024. PMID: 38250473 Free PMC article.
-
Involvement of the Voltage-Gated Calcium Channels L- P/Q- and N-Types in Synapse Elimination During Neuromuscular Junction Development.Mol Neurobiol. 2022 Jul;59(7):4044-4064. doi: 10.1007/s12035-022-02818-2. Epub 2022 Apr 27. Mol Neurobiol. 2022. PMID: 35474562 Free PMC article.
-
Purinergic Receptors in Neurological Diseases With Motor Symptoms: Targets for Therapy.Front Pharmacol. 2018 Apr 10;9:325. doi: 10.3389/fphar.2018.00325. eCollection 2018. Front Pharmacol. 2018. PMID: 29692728 Free PMC article. Review.
-
Membrane Receptor-Induced Changes of the Protein Kinases A and C Activity May Play a Leading Role in Promoting Developmental Synapse Elimination at the Neuromuscular Junction.Front Mol Neurosci. 2017 Aug 9;10:255. doi: 10.3389/fnmol.2017.00255. eCollection 2017. Front Mol Neurosci. 2017. PMID: 28848391 Free PMC article.
-
Tetanic depression is overcome by tonic adenosine A(2A) receptor facilitation of L-type Ca(2+) influx into rat motor nerve terminals.J Physiol. 2004 Oct 1;560(Pt 1):157-68. doi: 10.1113/jphysiol.2004.067595. Epub 2004 Aug 5. J Physiol. 2004. PMID: 15297571 Free PMC article.
References
-
- ANGLESON J.K., BETZ W.J. Intraterminal Ca2+ and spontaneous transmitter release at the frog neuromuscular junction. J. Neurophysiol. 2001;85:287–294. - PubMed
-
- BOWERSOX S.S., MILJANICH G.P., SUGIURA Y., LI C., NADASDI L., HOFFMAN B.B., RAMACHANDRAN J., KO C.P. Differential blockade of voltage-sensitive calcium channels at the mouse neuromuscular junction by novel omega-conopeptides and omega-agatoxin-IVA. J. Pharmacol. Exp. Ther. 1995;273:248–256. - PubMed
-
- CAPOGNA M., GÄHWILER B.H., THOMPSON S.M. Presynaptic inhibition of calcium-dependent and -independent release elicited with ionomycin, gadolinium, and α-latrotoxin in the hippocampus. J. Neurophysiol. 1996;75:2017–2028. - PubMed
-
- CUNHA R.A., SEBASTIÃO A.M. Extracellular metabolism of adenine nucleotides and adenosine in the innervated skeletal muscle of the frog. Eur. J. Pharmacol. 1991;197:83–92. - PubMed
-
- CUNHA R.A., SEBASTIÃO A.M. Adenosine and adenine nucleotides are independently released from both the nerve terminals and the muscle fibres upon electrical stimulation of the innervated skeletal muscle of the frog. Pflügers Arch. 1993;424:503–510. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
