

Dysregulated LIGHT expression on T cells mediates intestinal inflammation and contributes to IgA nephropathy
- PMID: 15067315
- PMCID: PMC362120
- DOI: 10.1172/JCI20096
Dysregulated LIGHT expression on T cells mediates intestinal inflammation and contributes to IgA nephropathy
Abstract
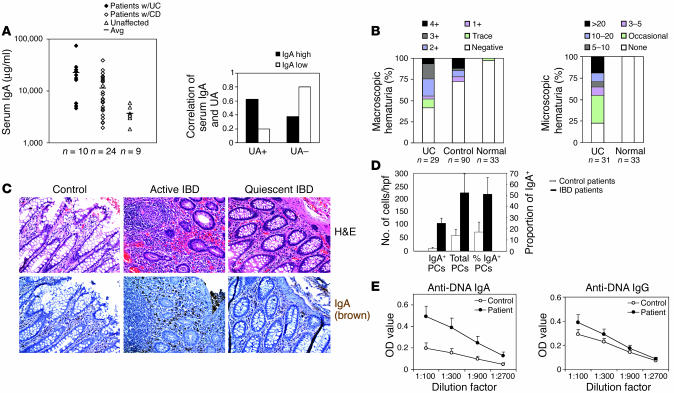
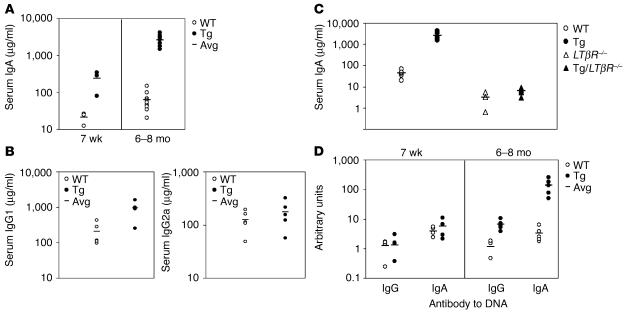
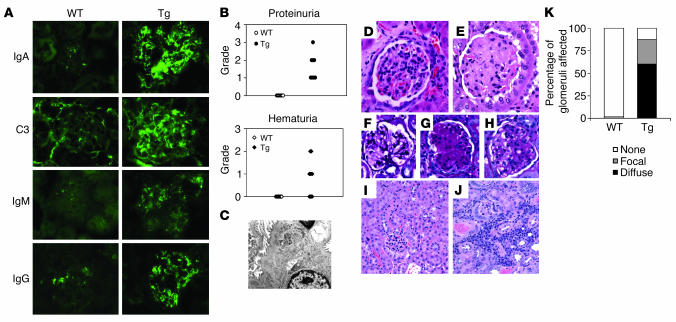
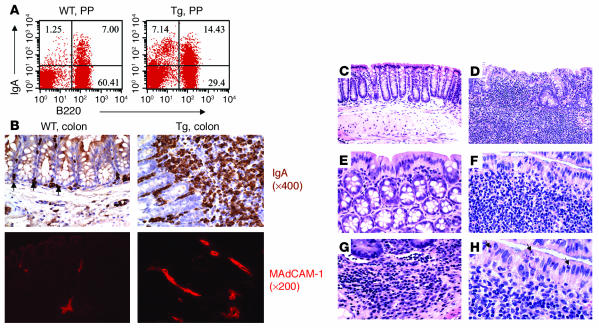
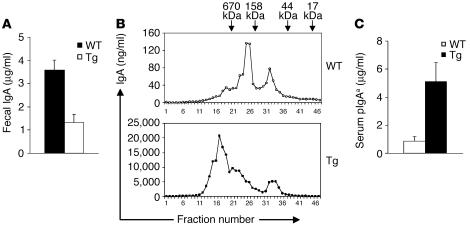
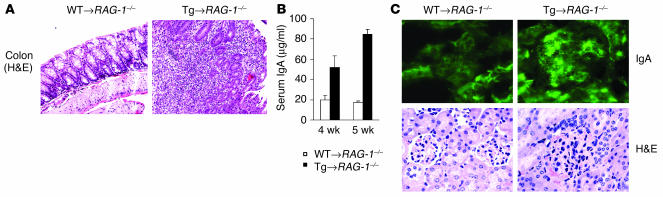
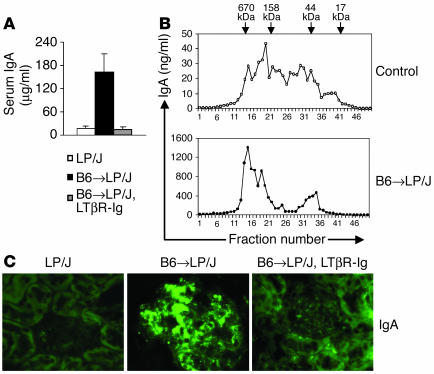
Whether and how T cells contribute to the pathogenesis of immunoglobulin A nephropathy (IgAN) has not been well defined. Here, we explore a murine model that spontaneously develops T cell-mediated intestinal inflammation accompanied by pathological features similar to those of human IgAN. Intestinal inflammation mediated by LIGHT, a ligand for lymphotoxin beta receptor (LTbetaR), not only stimulates IgA overproduction in the gut but also results in defective IgA transportation into the gut lumen, causing a dramatic increase in serum polymeric IgA. Engagement of LTbetaR by LIGHT is essential for both intestinal inflammation and hyperserum IgA syndrome in our LIGHT transgenic model. Impressively, the majority of patients with inflammatory bowel disease showed increased IgA-producing cells in the gut, elevated serum IgA levels, and severe hematuria, a hallmark of IgAN. These observations indicate the critical contributions of dysregulated LIGHT expression and intestinal inflammation to the pathogenesis of IgAN.
Figures
Similar articles
-
The Gut-Renal Connection in IgA Nephropathy.Semin Nephrol. 2018 Sep;38(5):504-512. doi: 10.1016/j.semnephrol.2018.05.020. Semin Nephrol. 2018. PMID: 30177022 Review.
-
Lymphtoxin beta receptor-Ig ameliorates TNBS-induced colitis via blocking LIGHT/HVEM signaling.Pharmacol Res. 2005 Sep;52(3):234-44. doi: 10.1016/j.phrs.2005.03.009. Pharmacol Res. 2005. PMID: 15925518
-
Constitutive expression of LIGHT on T cells leads to lymphocyte activation, inflammation, and tissue destruction.J Immunol. 2001 Dec 1;167(11):6330-7. doi: 10.4049/jimmunol.167.11.6330. J Immunol. 2001. PMID: 11714797
-
BAFF induces a hyper-IgA syndrome in the intestinal lamina propria concomitant with IgA deposition in the kidney independent of LIGHT.Cell Immunol. 2006 Jun;241(2):85-94. doi: 10.1016/j.cellimm.2006.08.002. Epub 2006 Sep 20. Cell Immunol. 2006. PMID: 16987502
-
Pathogenesis of IgA nephropathy.Ann Med Interne (Paris). 1999 Feb;150(2):91-8. Ann Med Interne (Paris). 1999. PMID: 10392257 Review.
Cited by
-
Mucosal Immune System Dysregulation in the Pathogenesis of IgA Nephropathy.Biomedicines. 2022 Nov 24;10(12):3027. doi: 10.3390/biomedicines10123027. Biomedicines. 2022. PMID: 36551783 Free PMC article. Review.
-
Title IgA Nephropathy and Oral Bacterial Species Related to Dental Caries and Periodontitis.Int J Mol Sci. 2022 Jan 10;23(2):725. doi: 10.3390/ijms23020725. Int J Mol Sci. 2022. PMID: 35054910 Free PMC article. Review.
-
LIGHT signals directly to intestinal epithelia to cause barrier dysfunction via cytoskeletal and endocytic mechanisms.Gastroenterology. 2007 Jun;132(7):2383-94. doi: 10.1053/j.gastro.2007.02.052. Epub 2007 Feb 27. Gastroenterology. 2007. PMID: 17570213 Free PMC article.
-
Different renal manifestations associated with very early onset pediatric inflammatory bowel disease: case report and review of literature.BMC Nephrol. 2021 Apr 22;22(1):146. doi: 10.1186/s12882-021-02358-2. BMC Nephrol. 2021. PMID: 33888087 Free PMC article. Review.
-
Increased proportion of follicular helper T cells is associated with B cell activation and disease severity in IgA nephropathy.Front Immunol. 2022 Aug 2;13:901465. doi: 10.3389/fimmu.2022.901465. eCollection 2022. Front Immunol. 2022. PMID: 35983053 Free PMC article.
References
-
- Gardby E, Lane P, Lycke NY. Requirements for B7-CD28 costimulation in mucosal IgA responses: paradoxes observed in CTLA4-H γ 1 transgenic mice. J. Immunol. 1998;161:49–59. - PubMed
-
- Vajdy M, Kosco-Vilbois MH, Kopf M, Kohler G, Lycke N. Impaired mucosal immune responses in interleukin 4-targeted mice. J. Exp. Med. 1995;181:41–53. - PubMed
-
- Hornquist CE, Ekman L, Grdic KD, Schon K, Lycke NY. Paradoxical IgA immunity in CD4-deficient mice. Lack of cholera toxin-specific protective immunity despite normal gut mucosal IgA differentiation. J. Immunol. 1995;155:2877–2887. - PubMed
-
- Lycke N, Eriksen L, Holmgren J. Protection against cholera toxin after oral immunization is thymus-dependent and associated with intestinal production of neutralizing IgA antitoxin. Scand. J. Immunol. 1987;25:413–419. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
