

Reduction of single-neuron firing uncertainty by cortical ensembles during motor skill learning
- PMID: 15071105
- PMCID: PMC6729737
- DOI: 10.1523/JNEUROSCI.5361-03.2004
Reduction of single-neuron firing uncertainty by cortical ensembles during motor skill learning
Abstract
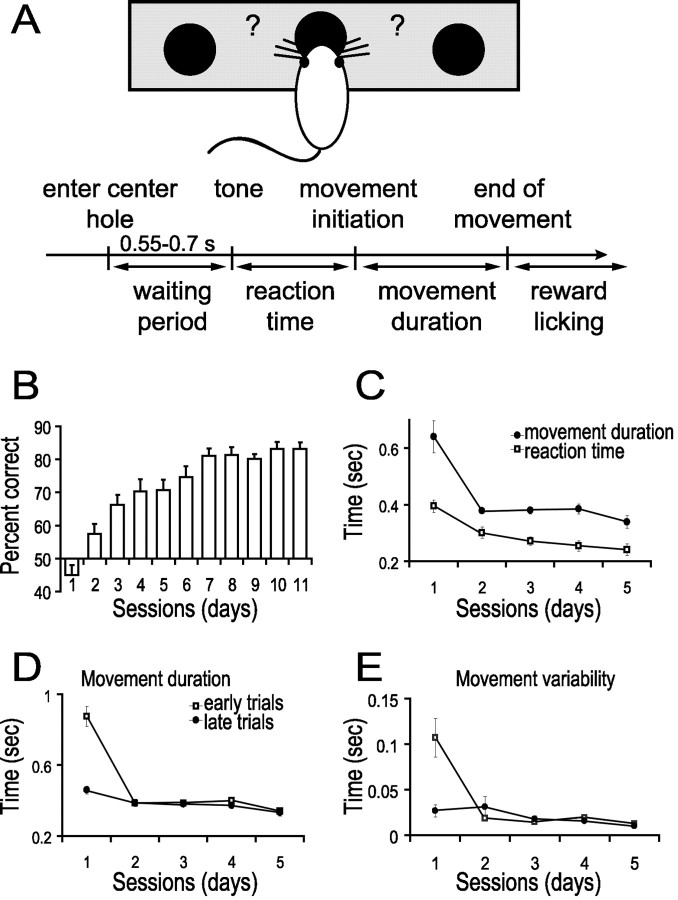
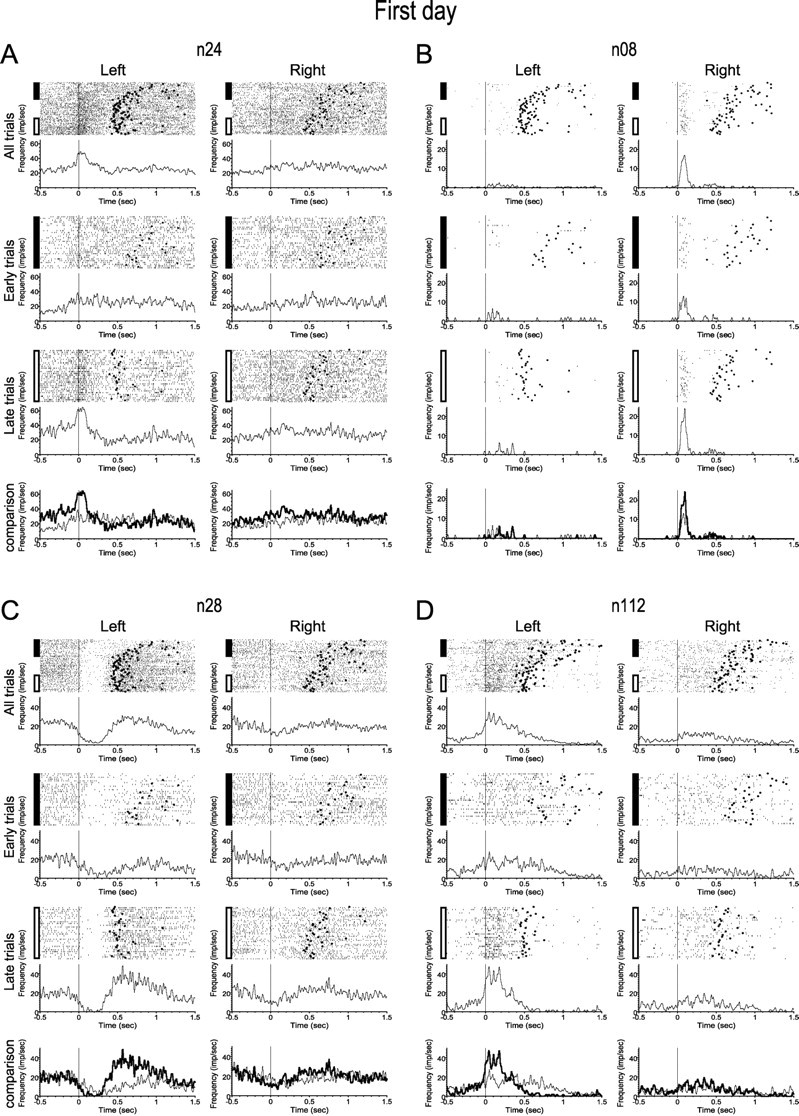
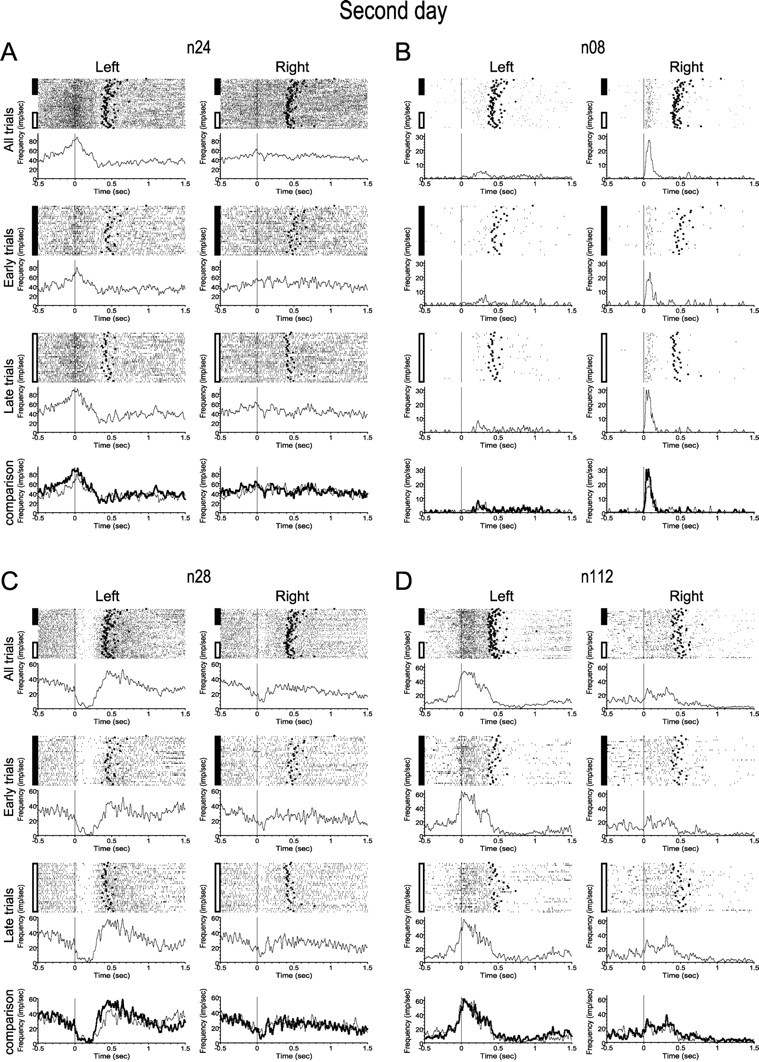
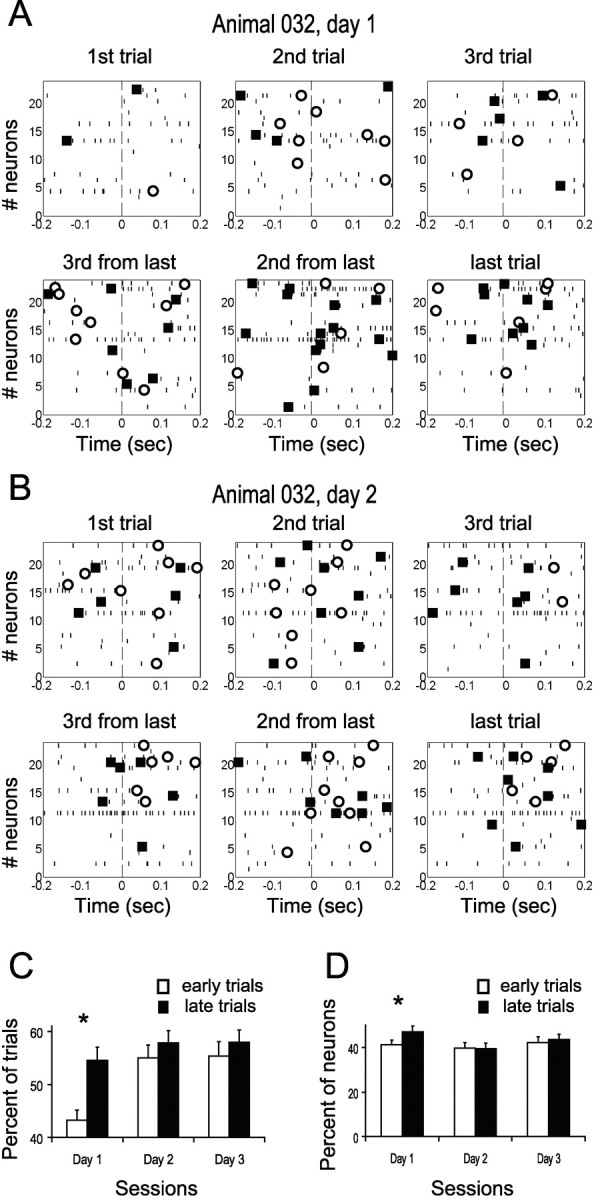
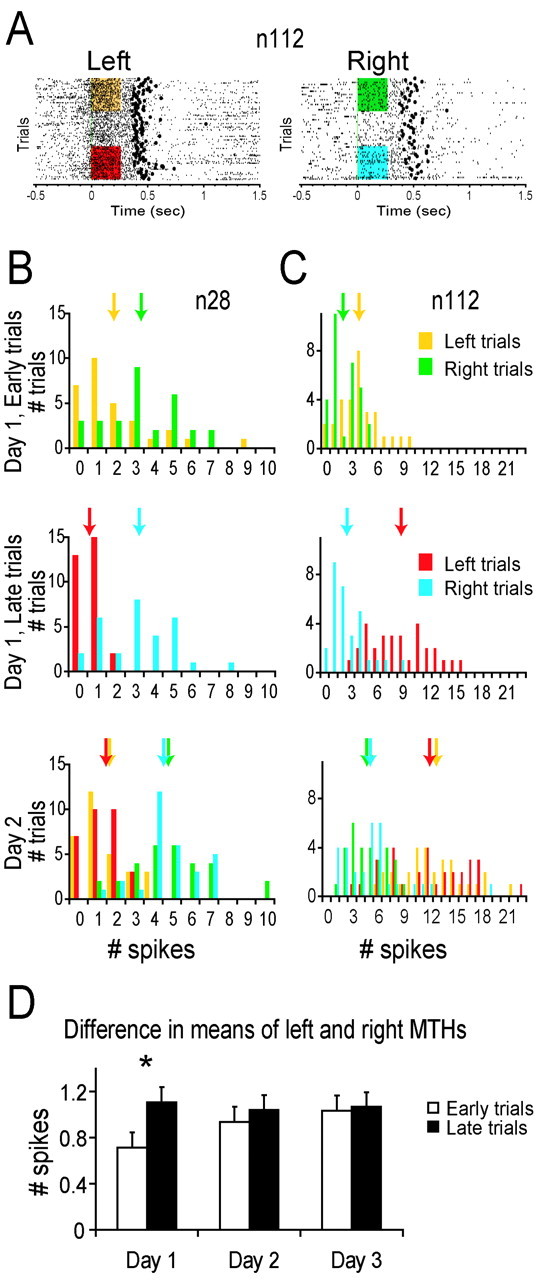
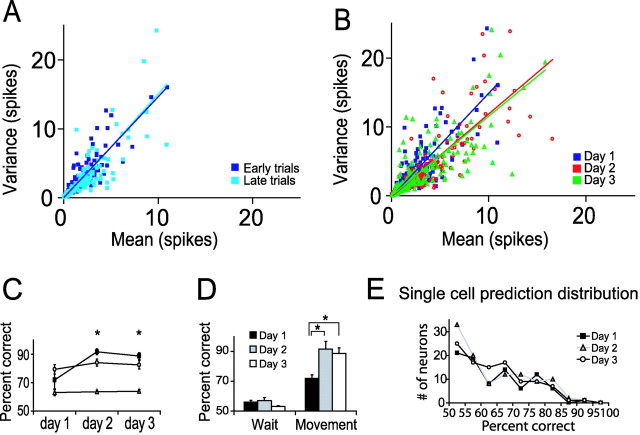
Motor skill learning is usually characterized by shortening of response time and performance of faster, more stereotypical movements. However, little is known about the changes in neural activity that underlie these behavioral changes. Here we used chronically implanted electrode arrays to record neuronal activity in the rat primary motor cortex (MI) as animals learned to execute movements in two directions. Strong modulation of MI single-neuron activity was observed while movement duration of the animal decreased. Despite many learning-induced changes, the precision with which single neurons fire did not improve with learning. Hence, prediction of movement direction from single neurons was bounded. In contrast, prediction of movement direction using neuronal ensembles improved significantly with learning, suggesting that, with practice, neuronal ensembles learn to overcome the uncertainty introduced by single-neuron stochastic activity.
Figures
References
-
- Bonato C, Zanette G, Fiaschi A, Rossini PM (2002) Activity-dependent modulation of synaptic transmission in the intact human motor cortex revealed with transcranial magnetic stimulation. Cereb Cortex 12: 1057–1062. - PubMed
-
- Dayan P, Abbott L (2001) Theoretical neuroscience: computational and mathematical modeling of neural systems. Cambridge, MA: MIT.
-
- Dubnov S, El-Yaniv R, Gdalyahu Y, Schneidman E, Tishby N, Yona G (2002) A new nonparametric clustering algorithm based on iterative estimation of distance profiles. Machine Learn 47: 35–61.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources