

Hgc1, a novel hypha-specific G1 cyclin-related protein regulates Candida albicans hyphal morphogenesis
- PMID: 15071502
- PMCID: PMC394249
- DOI: 10.1038/sj.emboj.7600195
Hgc1, a novel hypha-specific G1 cyclin-related protein regulates Candida albicans hyphal morphogenesis
Abstract
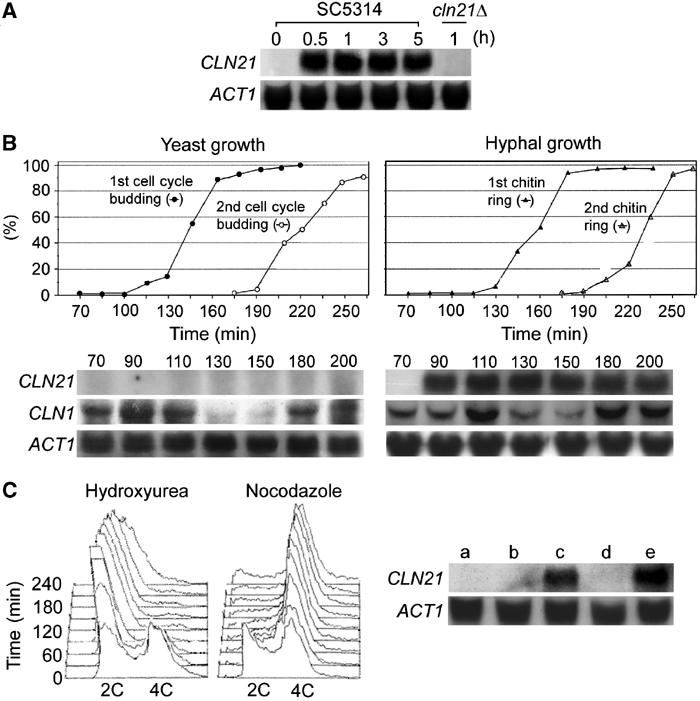
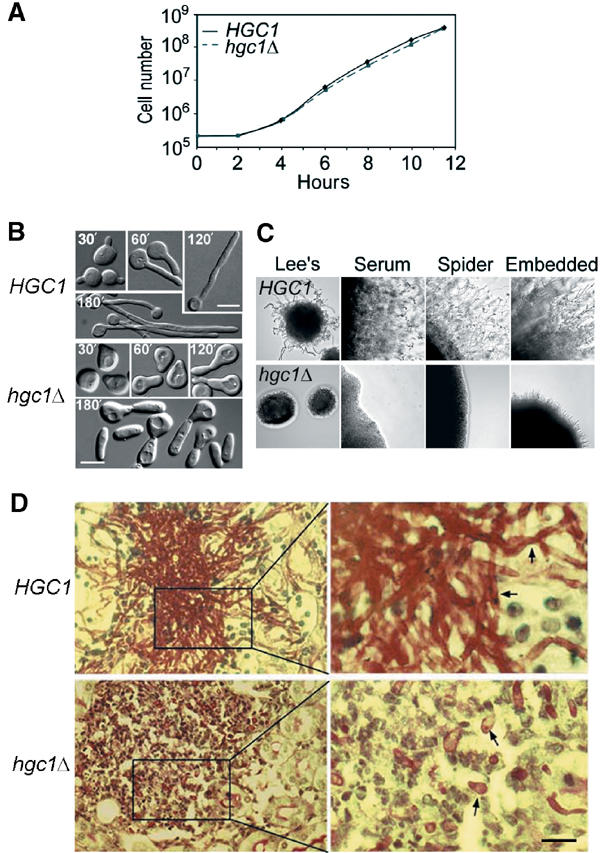
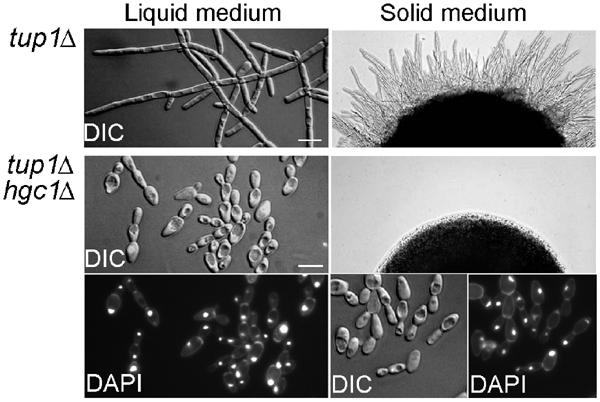
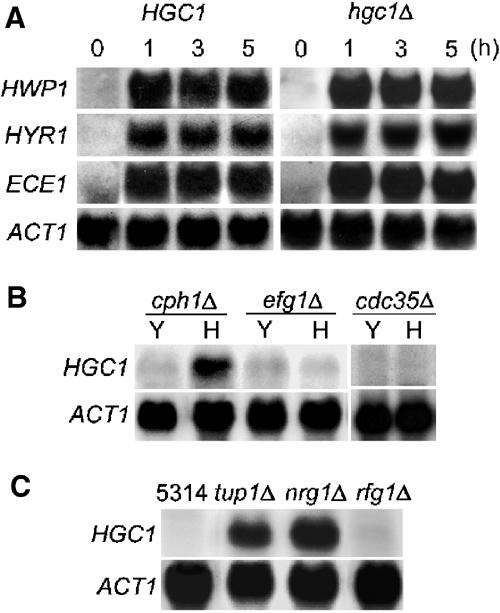
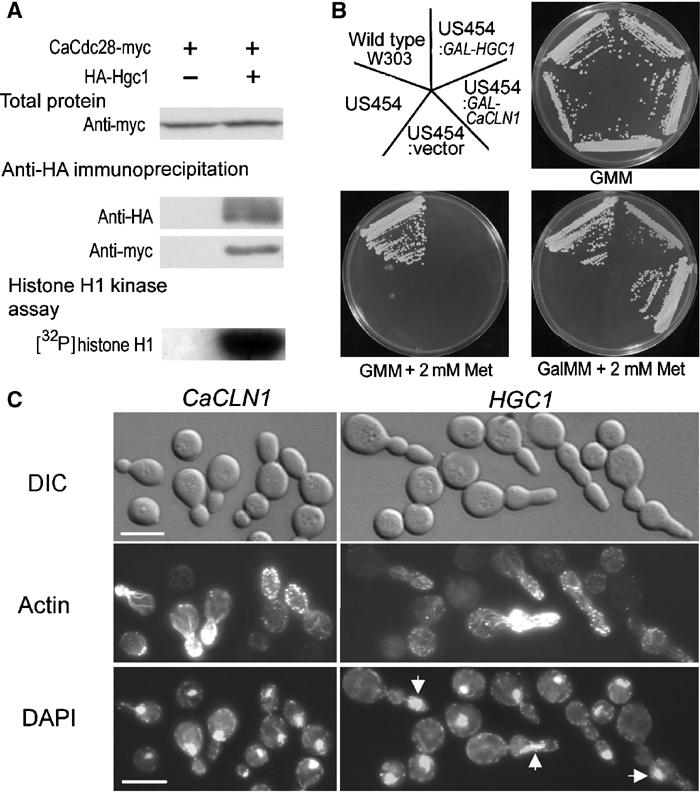
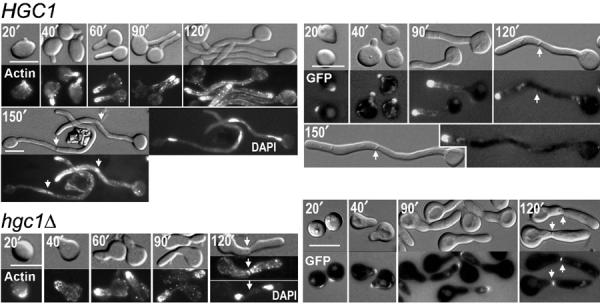
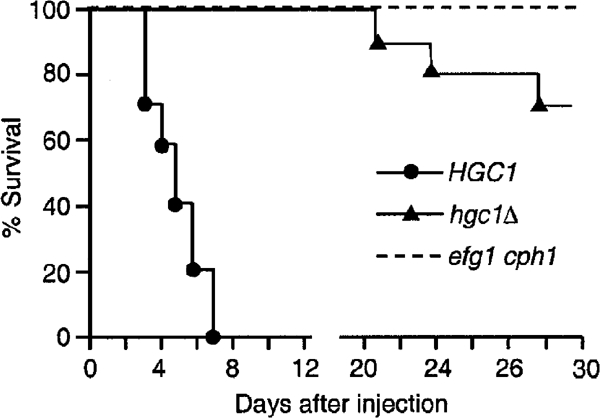
The human fungal pathogen Candida albicans switches from yeast to hyphal growth when exposed to serum or phagocytosed. However, the importance of this morphological switch for virulence remains highly controversial due to the lack of a mutant that affects hyphal morphogenesis only. Although many genes specifically expressed in hyphal cells have been identified and shown to encode virulence factors, none is required for hyphal morphogenesis. Here we report the first hypha-specific gene identified, HGC1, which is essential for hyphal morphogenesis. Deletion of HGC1 abolished hyphal growth in all laboratory conditions tested and in the kidneys of systemically infected mice with markedly reduced virulence. HGC1 expression is co-regulated with other virulence genes such as HWP1 by the cAMP/protein kinase A signaling pathway and transcriptional repressor Tup1/Nrg1. Hgc1 is a G1 cyclin-related protein and co-precipitated with the cyclin-dependent kinase (Cdk) CaCdc28. It has recently emerged that cyclin/Cdk complexes promote other forms of polarized cell growth such as tumor cell migration and neurite outgrowth. C. albicans seems to have adapted a conserved strategy to control specifically hyphal morphogenesis.
Figures

References
-
- Andrews B, Measday V (1998) The cyclin family of budding yeast: abundant use of a good idea. Trends Genet 14: 66–72 - PubMed
-
- Bai C, Ramanan N, Wang YM, Wang Y (2002) Spindle assembly checkpoint component CaMad2p is indispensable for Candida albicans survival and virulence in mice. Mol Microbiol 45: 31–44 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
