

Defects in the N-linked oligosaccharide biosynthetic pathway in a Trypanosoma brucei glycosylation mutant
- PMID: 15075256
- PMCID: PMC387663
- DOI: 10.1128/EC.3.2.255-263.2004
Defects in the N-linked oligosaccharide biosynthetic pathway in a Trypanosoma brucei glycosylation mutant
Abstract
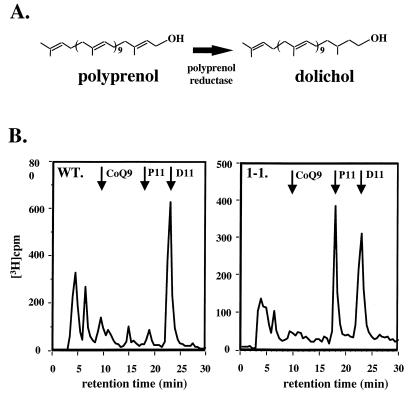
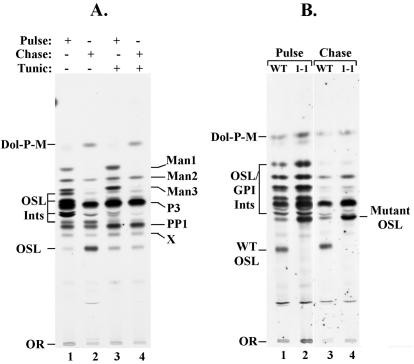
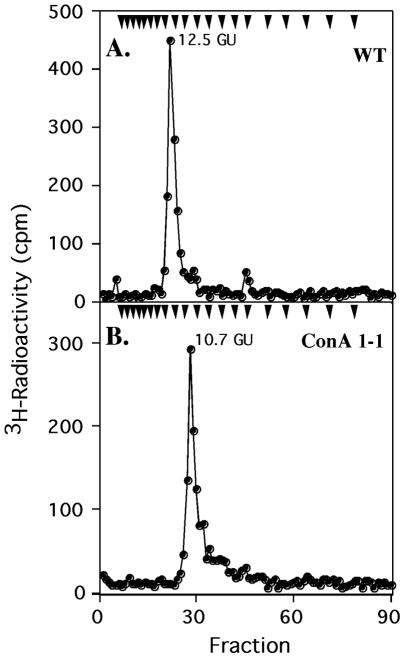
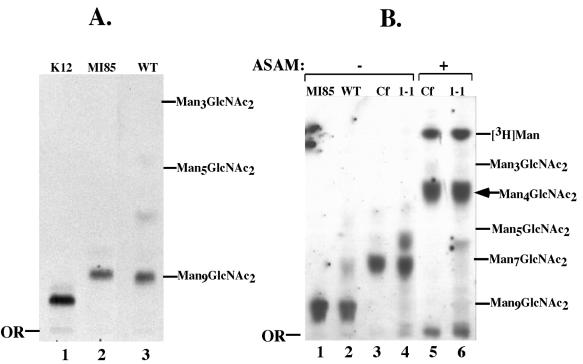
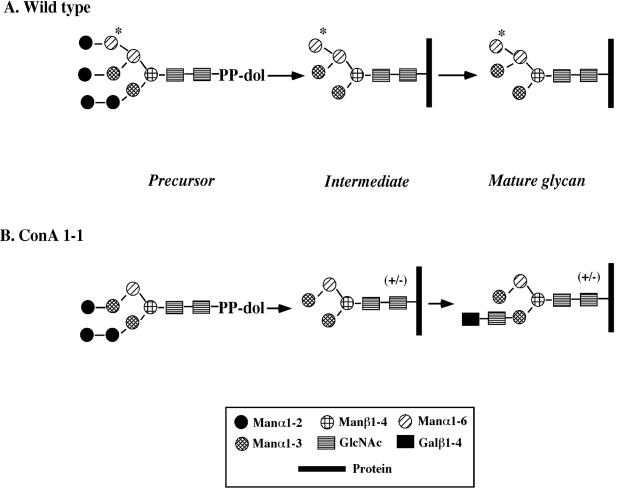
Concanavalin A (ConA) kills the procyclic (insect) form of Trypanosoma brucei by binding to its major surface glycoprotein, procyclin. We previously isolated a mutant cell line, ConA 1-1, that is less agglutinated and more resistant to ConA killing than are wild-type (WT) cells. Subsequently we found that the ConA resistance phenotype in this mutant is due to the fact that the procyclin either has no N-glycan or has an N-glycan with an altered structure. Here we demonstrate that the alteration in procyclin N-glycosylation correlates with two defects in the N-linked oligosaccharide biosynthetic pathway. First, ConA 1-1 has a defect in activity of polyprenol reductase, an enzyme involved in synthesis of dolichol. Metabolic incorporation of [3H]mevalonate showed that ConA 1-1 synthesizes equal amounts of dolichol and polyprenol, whereas WT cells make predominantly dolichol. Second, we found that ConA 1-1 synthesizes and accumulates an oligosaccharide lipid (OSL) precursor that is smaller in size than that from WT cells. The glycan of OSL in WT cells is apparently Man9GlcNAc2, whereas that from ConA 1-1 is Man7GlcNAc2. The smaller OSL glycan in the ConA 1-1 explains how some procyclin polypeptides bear a Man4GlcNAc2 modified with a terminal N-acetyllactosamine group, which is poorly recognized by ConA.
Figures

References
-
- Acosta-Serrano, A., R. N. Cole, and P. T. Englund. 2000. Killing of Trypanosoma brucei by concanavalin A: structural basis of resistance in glycosylation mutants. J. Mol. Biol. 304:633-644. - PubMed
-
- Acosta-Serrano, A., R. N. Cole, A. Mehlert, M. G. Lee, M. A. Ferguson, and P. T. Englund. 1999. The procyclin repertoire of Trypanosoma brucei. Identification and structural characterization of the Glu-Pro-rich polypeptides. J. Biol. Chem. 274:29763-29771. - PubMed
-
- Brun, R., and M. Shonenberger. 1979. Cultivation and in vitro cloning of procyclic culture forms of Trypanosoma brucei in a semi-defined medium. Acta Trop. 36:289-292. - PubMed
-
- Clayton, C. E., and M. R. Mowatt. 1989. The procyclic acidic repetitive proteins of Trypanosoma brucei. Purification and post-translational modification. J. Biol. Chem. 264:15088-15093. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
