

LaeA, a regulator of secondary metabolism in Aspergillus spp
- PMID: 15075281
- PMCID: PMC387652
- DOI: 10.1128/EC.3.2.527-535.2004
LaeA, a regulator of secondary metabolism in Aspergillus spp
Abstract
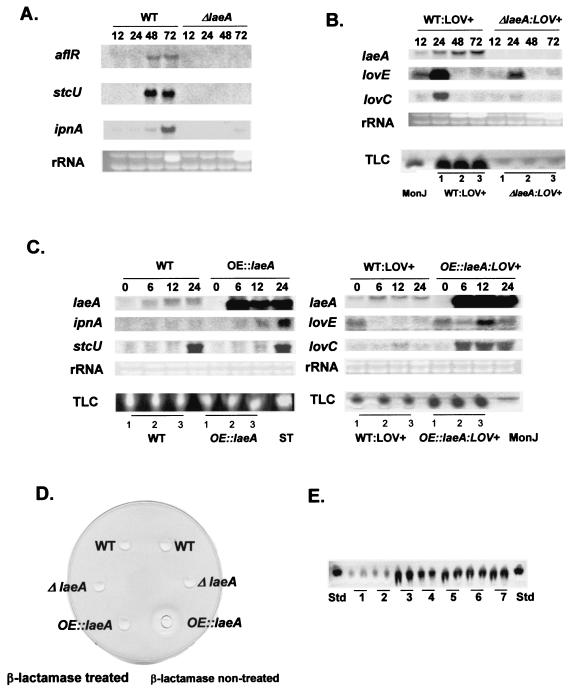
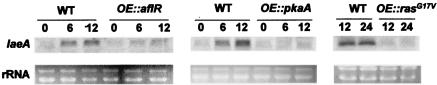
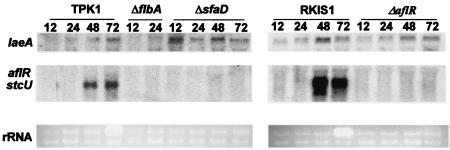
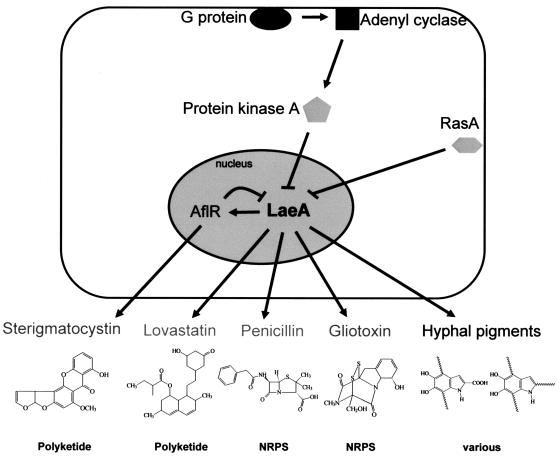
Secondary metabolites, or biochemical indicators of fungal development, are of intense interest to humankind due to their pharmaceutical and/or toxic properties. We present here a novel Aspergillus nuclear protein, LaeA, as a global regulator of secondary metabolism in this genus. Deletion of laeA (DeltalaeA) blocks the expression of metabolic gene clusters, including the sterigmatocystin (carcinogen), penicillin (antibiotic), and lovastatin (antihypercholesterolemic agent) gene clusters. Conversely, overexpression of laeA triggers increased penicillin and lovastatin gene transcription and subsequent product formation. laeA expression is negatively regulated by AflR, a sterigmatocystin Zn2Cys6 transcription factor, in a unique feedback loop, as well as by two signal transduction elements, protein kinase A and RasA. Although these last two proteins also negatively regulate sporulation, DeltalaeA strains show little difference in spore production compared to the wild type, indicating that the primary role of LaeA is to regulate metabolic gene clusters.
Figures
References
-
- Adams, T. H., and J. H. Yu. 1998. Coordinate control of secondary metabolite production and asexual sporulation in Aspergillus nidulans. Curr. Opin. Microbiol. 1:674-677. - PubMed
-
- Belkacemi, L., R. C. Barton, V. Hopwood, and E. G. V. Evans. 1999. Determination of optimum growth conditions for gliotoxin production by Aspergillus fumigatus and development of a novel method for gliotoxin detection. Med. Mycol. 37:227-233. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
