

doi: 10.1110/ps.03587604.
Epub 2004 Apr 9.
Critical nucleation size in the folding of small apparently two-state proteins
Affiliations
- PMID: 15075405
- PMCID: PMC2286761
- DOI: 10.1110/ps.03587604
Item in Clipboard
Critical nucleation size in the folding of small apparently two-state proteins
Protein Sci.
2004 May.
Abstract
For apparently two-state proteins, we found that the size (number of folded residues) of a transition state is mostly encoded by the topology, defined by total contact distance (TCD) of the native state, and correlates with its folding rate. This is demonstrated by using a simple procedure to reduce the native structures of the 41 two-state proteins with native TCD as a constraint, and is further supported by analyzing the results of eight proteins from protein engineering studies. These results support the hypothesis that the major rate-limiting process in the folding of small apparently two-state proteins is the search for a critical number of residues with the topology close to that of the native state.
Figures
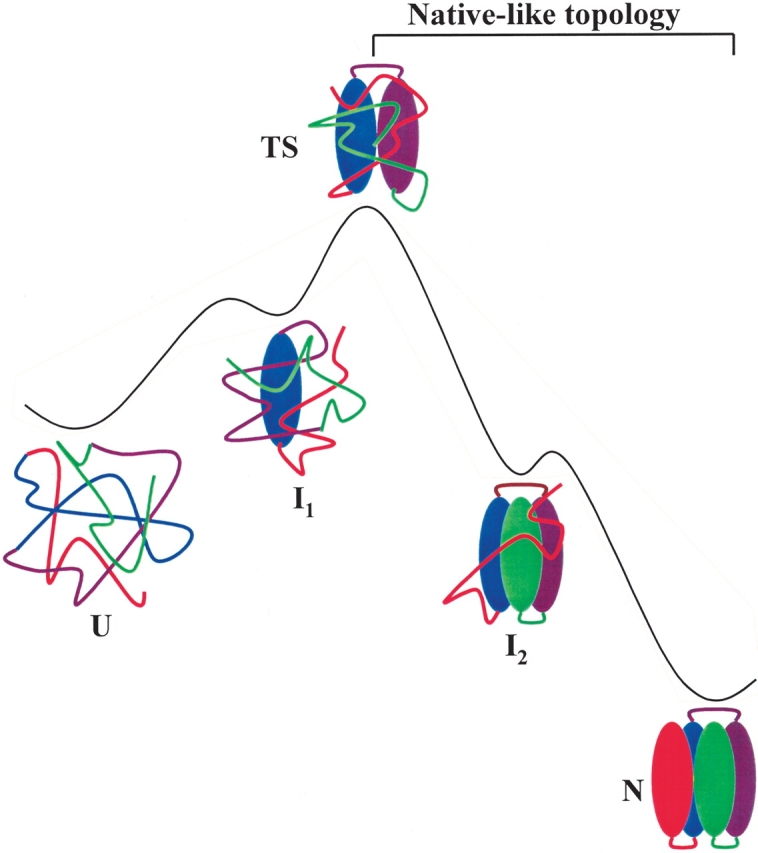
A schematic diagram of a putative folding pathway for an apparent two-state protein and the change of topology along the folding coordinate. In this illustration, the formation of the middle region is the rate-determining step. The two hidden intermediates are not observable in the conventional kinetic folding experiments. I2 can be identified by native-state hydrogen exchange method (Chu et al. 2002).
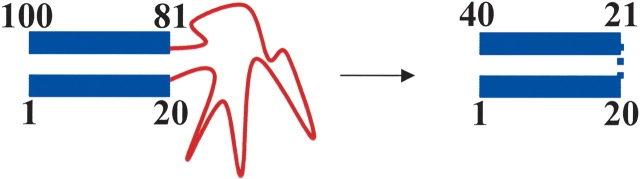
Illustration for calculation of TCD of a partially unfolded structure. The unfolded region was substituted with a putative bond without sequence separation. In this illustration, the C-terminal region following the unfolded loop was renumbered after the loop was substituted with a putative bind. The TCD value was calculated using the structured region with renumbered sequence to obtain |i - j| values in equation 1.
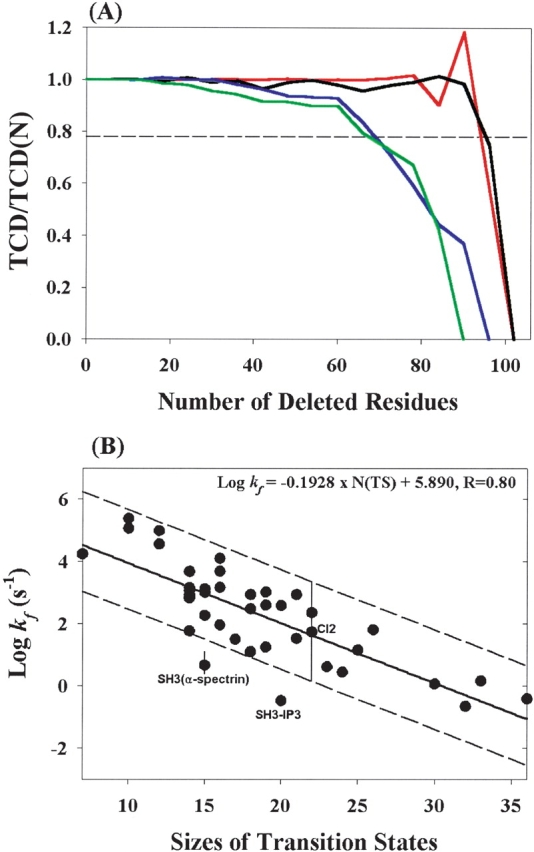
Reduction of the native structures. (A) Plots of TCD/TCD(N) vs. the number of deleted residues for cyt. c (red, TCD(N) =0.84), Rd-apocyt b562 (blue, TCD(N) =0.79), mAcP (pink, TCD(N) =1.52), and TNfn3 (green, TCD(N) =1.24). TCD is the corresponding topology of the structure after deletion of a six-residue segment in each step (see Materials and Methods) and TCD(N) is the topology for the native state structure. The dashed line represents 78% of the TCD(N) value. (B) Correlation between folding rates and the sizes of the “transition states” generated using the reduction procedures. The solid line is the result of a linear fitting. The dotted line indicates a ~50-fold deviation from the solid line determined by the maximum range of the folding rates from single mutants of CI2 (Jackson 1998). Note: The molecule (SH3–PI3) in the experiment had additional four residues (WNSS) at the C terminus (Guijarro et al. 1998) of the structure in the PDB file (2PNI) that is used to calculate the TCD value.
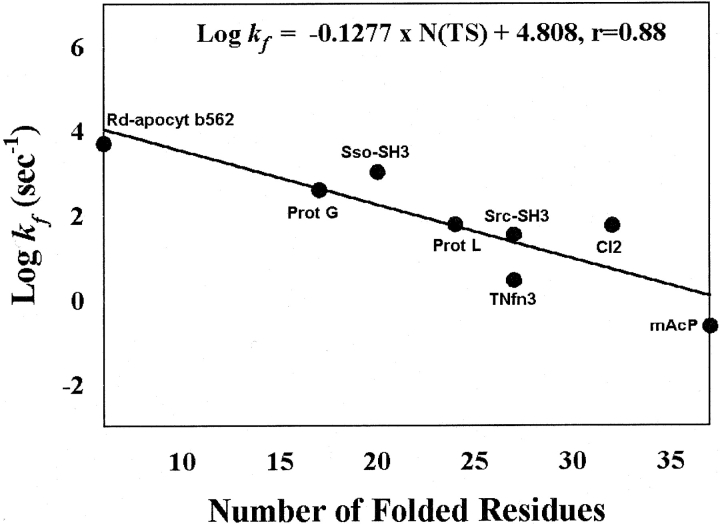
The correlation between the folding rate and the size of the transition state derived based on the experimental φ values (see Materials and Methods).
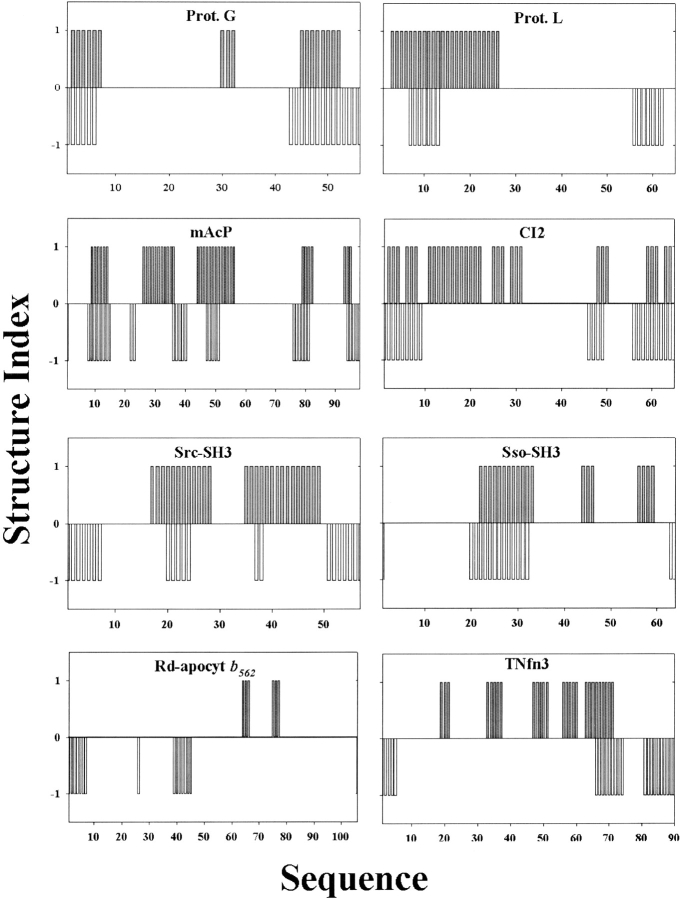
Comparison of the structures of transition states determined from experimental φ values and from the reduction procedure using structure indexes for the eight proteins. A residue or its nearest neighbor with φ > 0.35 is considered structured and given a structure index 1. Otherwise, it is given a structural index 0 to represent that the residue is unfolded in the transition state. Similarly, a residue that is folded in the transition state based on the reduction procedure is given a structure index of −1 (the minus sign is for the purpose of comparison). It is given a structural index of 0 if the residue is unfolded in the transition state.
References
-
- Alm, E., Morozov, A.V., Kortemme, T., and Baker, D. 2002. Simple physical models connect theory and experiment in protein folding kinetics. J. Mol.Biol. 322 463–476. - PubMed
-
- Baldwin, R.L. and Rose, G.D. 1999. Is protein folding hierarchic? I. Local structure and peptide folding. Trends Biochem. Sci. 24 26–33. - PubMed
-
- Chiti, F., Taddei, N., White, P.M., Bucciantini, M., Magherini, F., Stefani, M., and Dobson, C.M. 1999. Mutational analysis of acylphosphatase suggests the importance of topology and contact order in protein folding. Nat. Struct. Biol. 6 1005–1009. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
