

Misfolded proteins are sorted by a sequential checkpoint mechanism of ER quality control
- PMID: 15078901
- PMCID: PMC2172089
- DOI: 10.1083/jcb.200309132
Misfolded proteins are sorted by a sequential checkpoint mechanism of ER quality control
Abstract
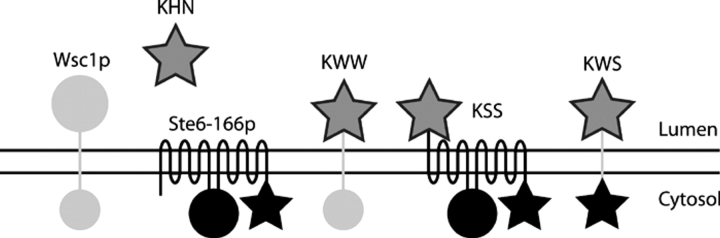
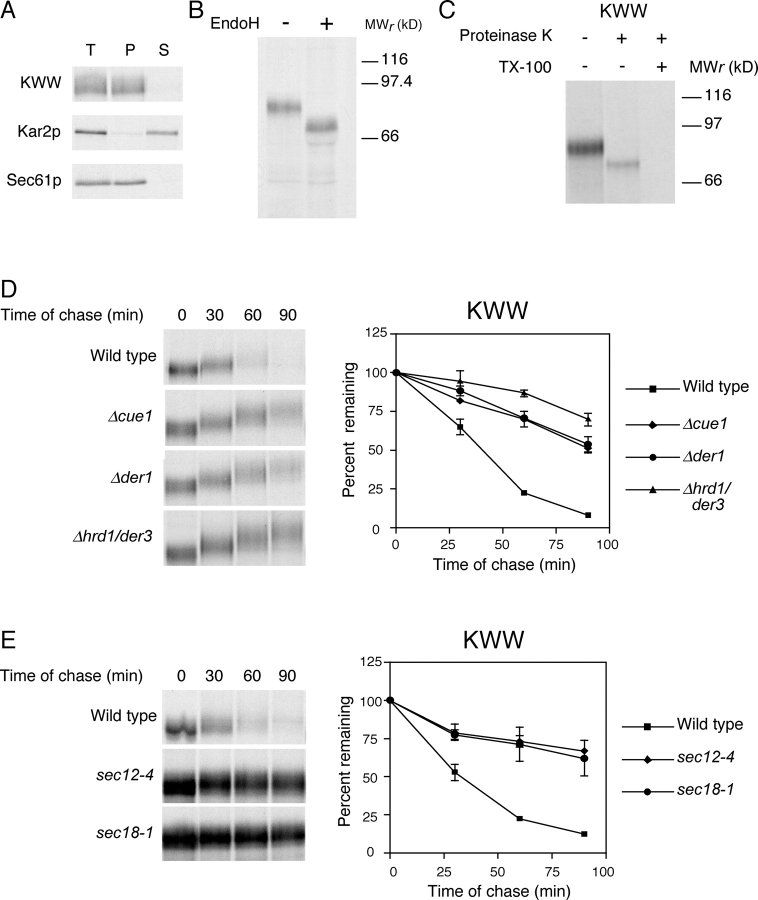
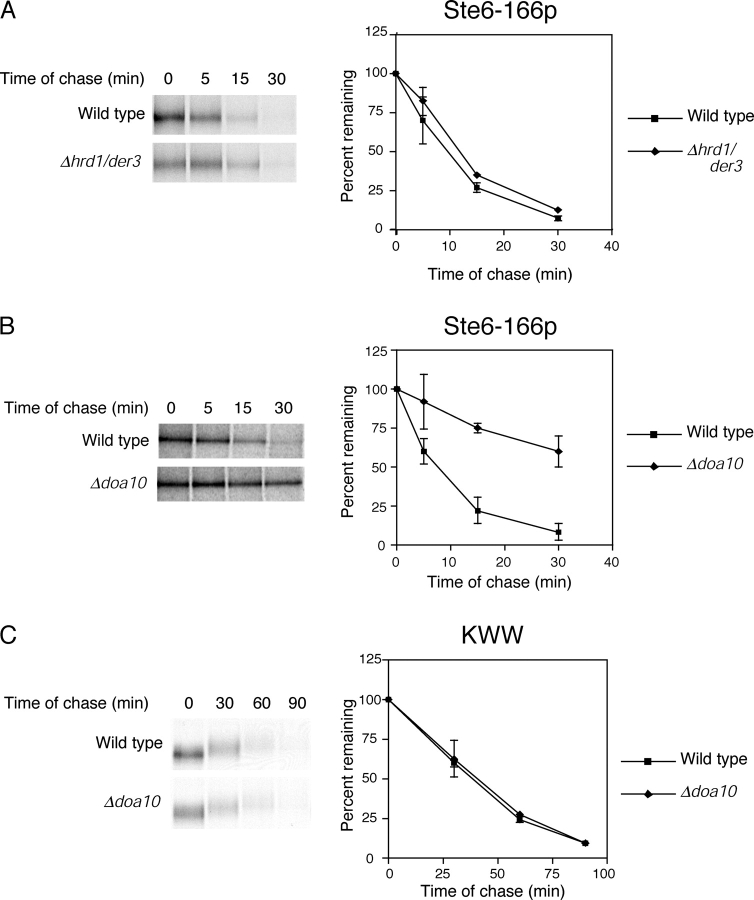
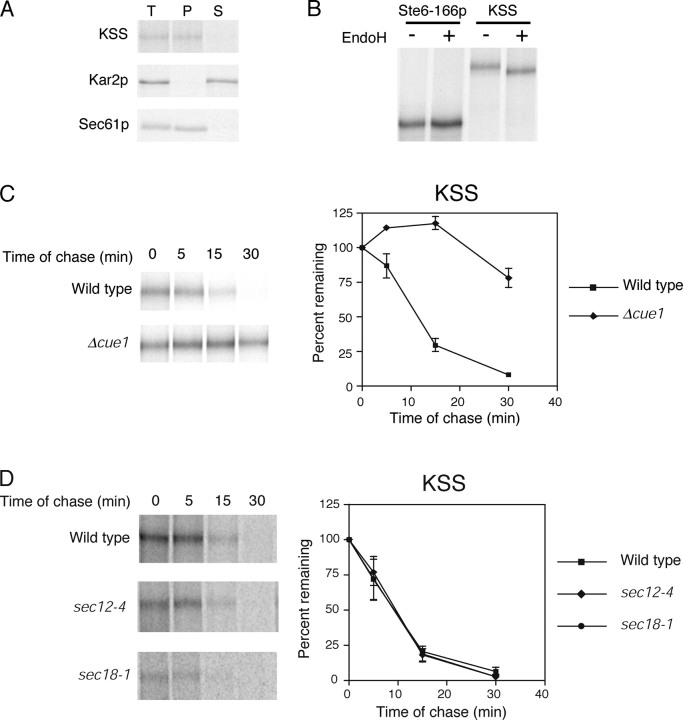
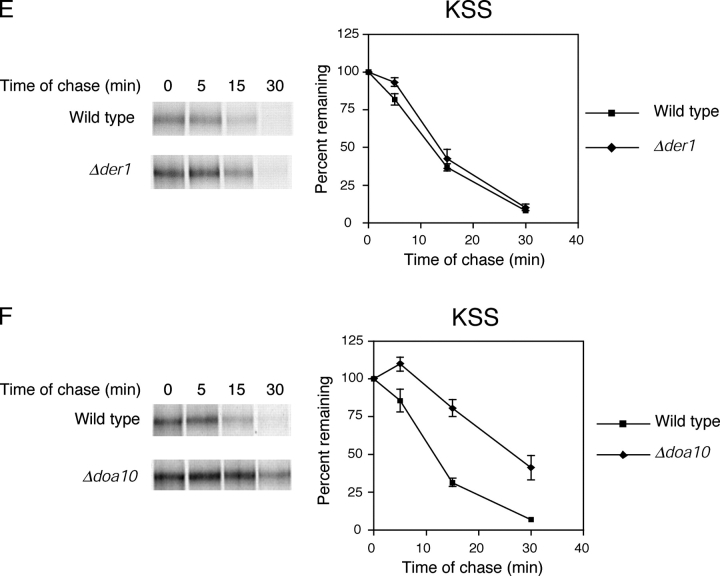
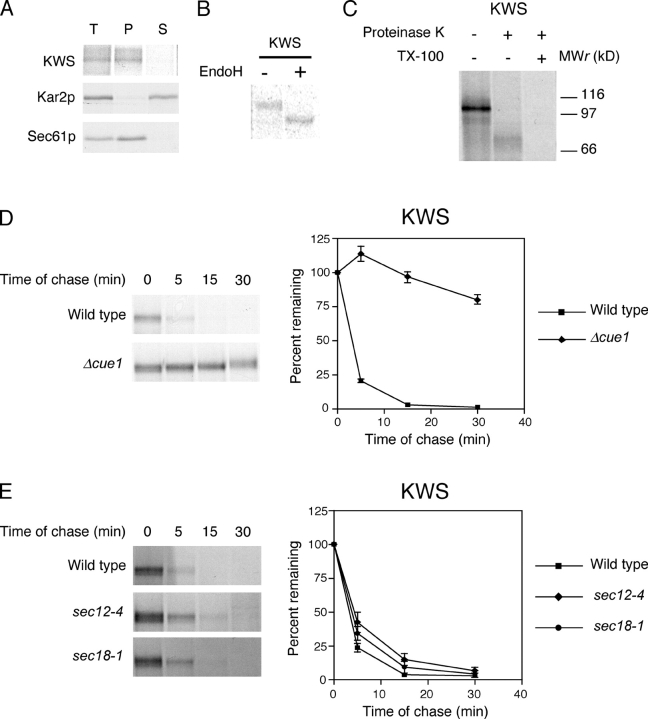
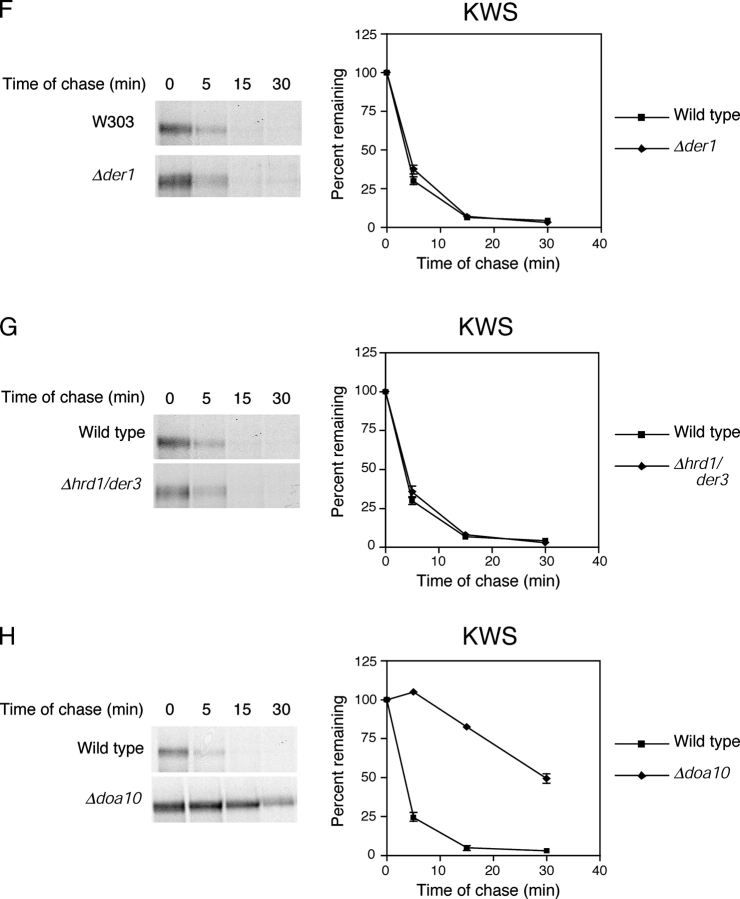
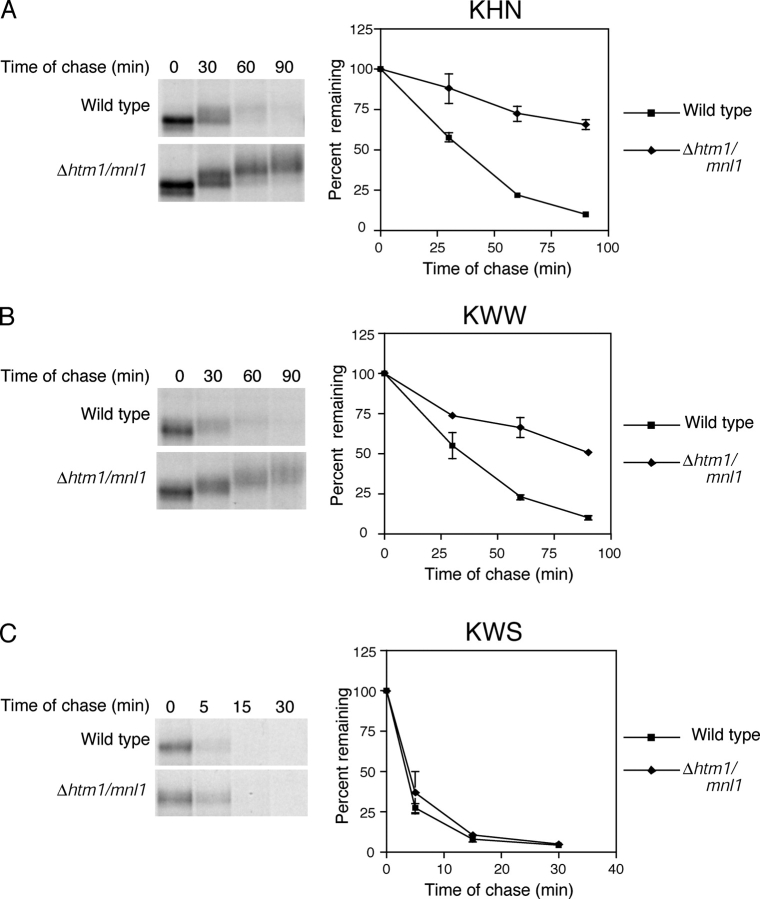
Misfolded proteins retained in the endoplasmic reticulum (ER) are degraded by the ER-associated degradation pathway. The mechanisms used to sort them from correctly folded proteins remain unclear. Analysis of substrates with defined folded and misfolded domains has revealed a system of sequential checkpoints that recognize topologically distinct domains of polypeptides. The first checkpoint examines the cytoplasmic domains of membrane proteins. If a lesion is detected, it is retained statically in the ER and rapidly degraded without regard to the state of its other domains. Proteins passing this test face a second checkpoint that monitors domains localized in the ER lumen. Proteins detected by this pathway are sorted from folded proteins and degraded by a quality control mechanism that requires ER-to-Golgi transport. Although the first checkpoint is obligatorily directed at membrane proteins, the second monitors both soluble and membrane proteins. Our data support a model whereby "properly folded" proteins are defined biologically as survivors that endure a series of distinct checkpoints.
Figures
References
-
- Arvan, P., X. Zhao, J. Ramos-Castaneda, and A. Chang. 2002. Secretory pathway quality control operating in Golgi, plasmalemmal, and endosomal systems. Traffic. 3:771–780. - PubMed
-
- Barlowe, C., and R. Schekman. 1993. SEC12 encodes a guanine-nucleotide-exchange factor essential for transport vesicle budding from the ER. Nature. 365:347–349. - PubMed
-
- Bays, N.W., R.G. Gardner, L.P. Seelig, C.A. Joazeiro, and R.Y. Hampton. 2001. Hrd1p/Der3p is a membrane-anchored ubiquitin ligase required for ER- associated degradation. Nat. Cell Biol. 3:24–29. - PubMed
-
- Belden, W.J., and C. Barlowe. 2001. Role of Erv29p in collecting soluble secretory proteins into ER-derived transport vesicles. Science. 294:1528–1531. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
