

Distinct aerobic and hypoxic mechanisms of HIF-alpha regulation by CSN5
- PMID: 15082527
- PMCID: PMC387414
- DOI: 10.1101/gad.1180104
Distinct aerobic and hypoxic mechanisms of HIF-alpha regulation by CSN5
Abstract
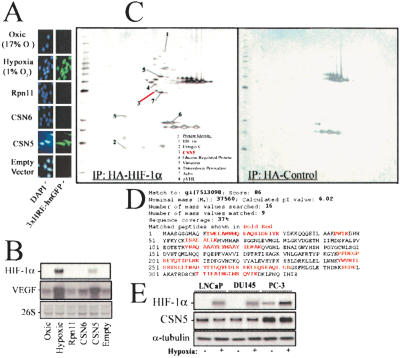
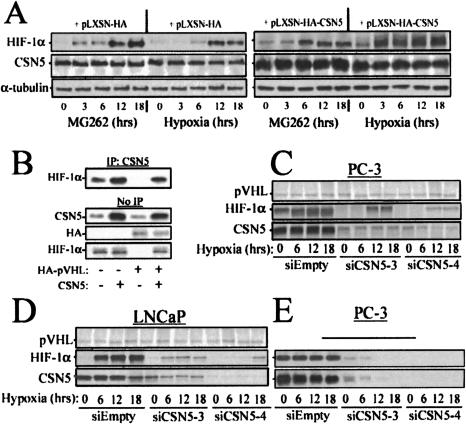
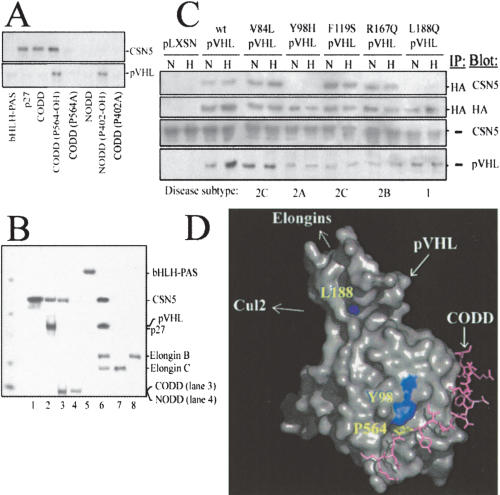
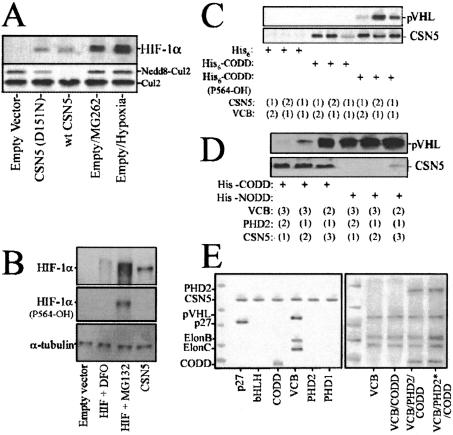
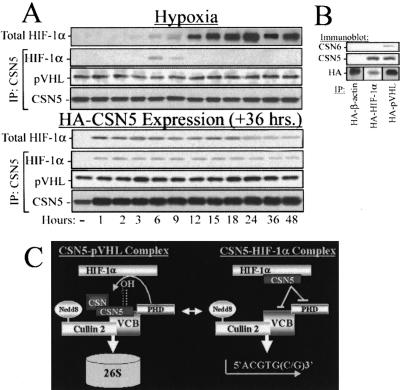
Mammalian oxygen homeostasis is dependent on the HIF family of transcription factors. The CSN subunit, CSN5, binds both the CODD of HIF-1 alpha and the pVHL tumor suppressor. High CSN5 expression generates a pVHL-independent form of CSN5 that stabilizes HIF-1 alpha aerobically by inhibiting HIF-1 alpha prolyl-564 hydroxylation. Aerobic CSN5 association with HIF-1 alpha occurs independently of the CSN holocomplex, leading to HIF-1 alpha stabilization independent of Cullin 2 deneddylation. CSN5 weakly associates with HIF-1 alpha under hypoxia, but is required for optimal hypoxia-mediated HIF-1 alpha stabilization. These results indicate that CSN5 regulates aerobic as well as hypoxic HIF-1 alpha stability by different mechanisms during oncogenesis.
Figures
References
-
- Bae M.K., Ahn, M.Y., Jeong, J.W., Bae, M.H., Lee, Y.M., Bae, S.K., Park, J.W., Kim, K.R., and Kim, K.W. 2002. Jab1 interacts directly with HIF-1alpha and regulates its stability. J. Biol. Chem. 277: 9-12. - PubMed
-
- Chan D.A., Sutphin, P.D., Denko, N.C., and Giaccia, A.J. 2002. Role of prolyl hydroxylation in oncogenically stabilized hypoxia-inducible factor-1alpha. J. Biol. Chem. 277: 40112-40117. - PubMed
-
- Denko N.C., Fontana, L.A., Hudson, K.M., Sutphin, P.D., Raychaudhuri, S., Altman, R., and Giaccia, A.J. 2003. Investigating hypoxic tumor physiology through gene expression patterns. Oncogene 22: 5907-5914. - PubMed
-
- Hoffman M.A., Ohh, M., Yang, H., Klco, J.M., Ivan, M., and Kaelin Jr., W.G. 2001. von Hippel-Lindau protein mutants linked to type 2C VHL disease preserve the ability to downregulate HIF. Hum. Mol. Genet. 10: 1019-1027. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases