

The ascorbic acid redox state controls guard cell signaling and stomatal movement
- PMID: 15084716
- PMCID: PMC423206
- DOI: 10.1105/tpc.021584
The ascorbic acid redox state controls guard cell signaling and stomatal movement
Abstract
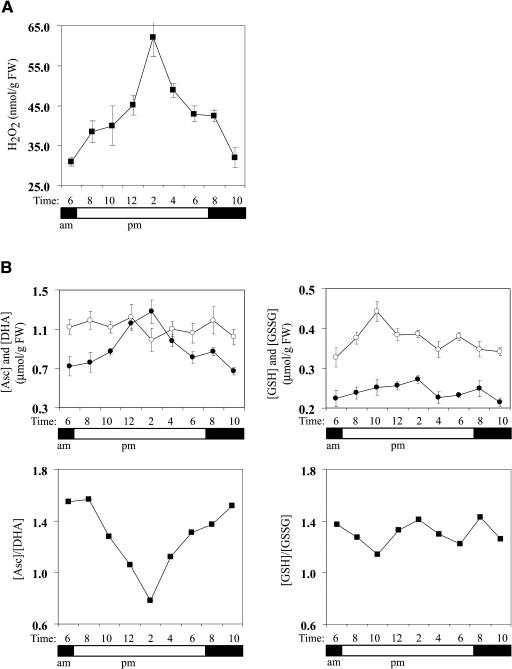
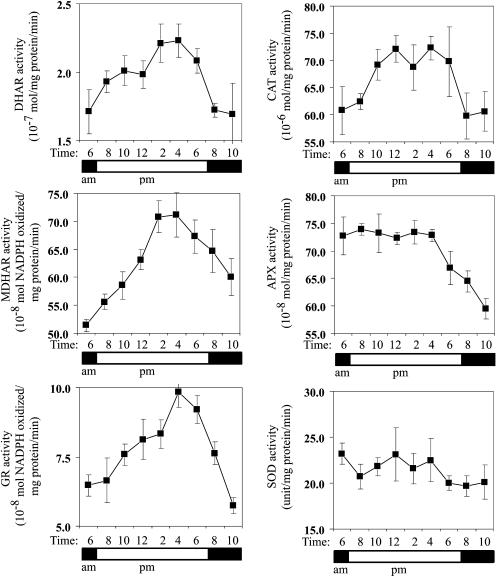
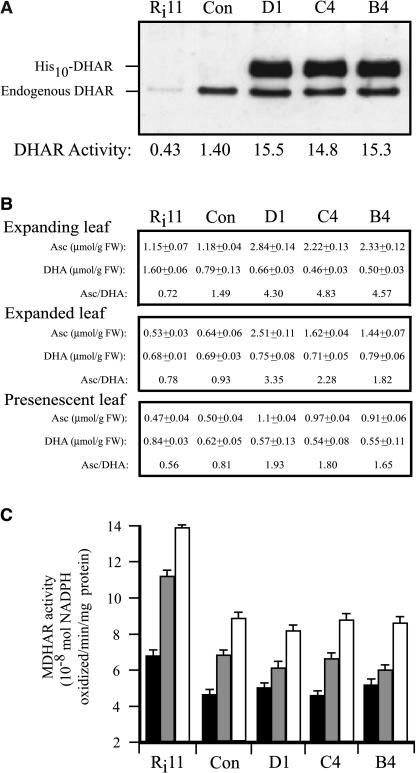
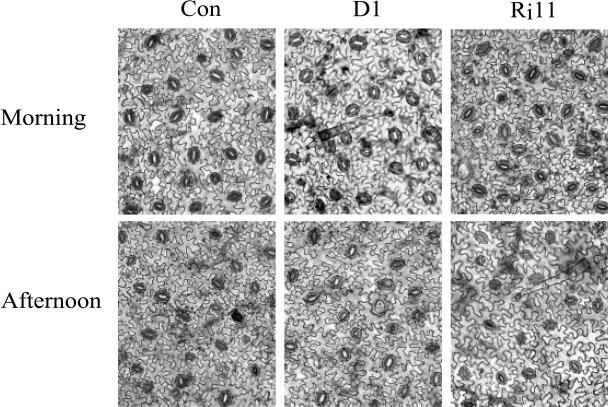
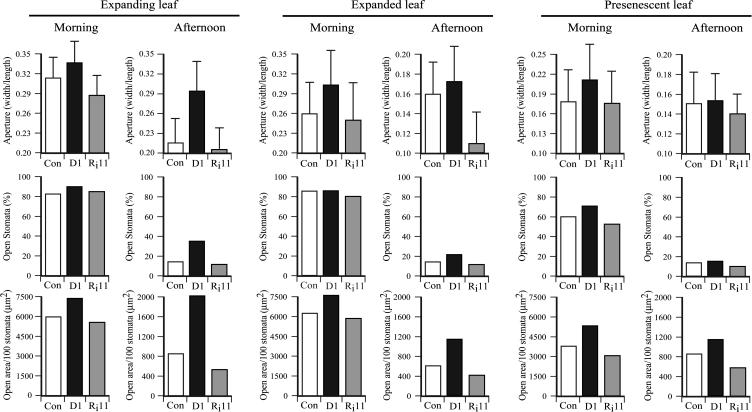
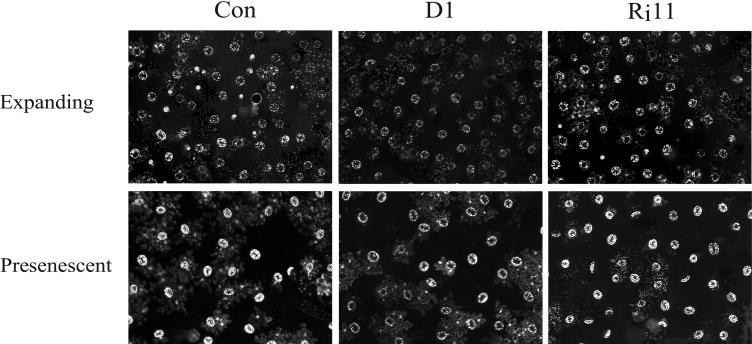
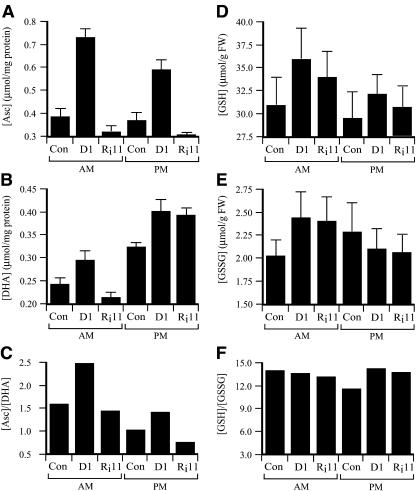
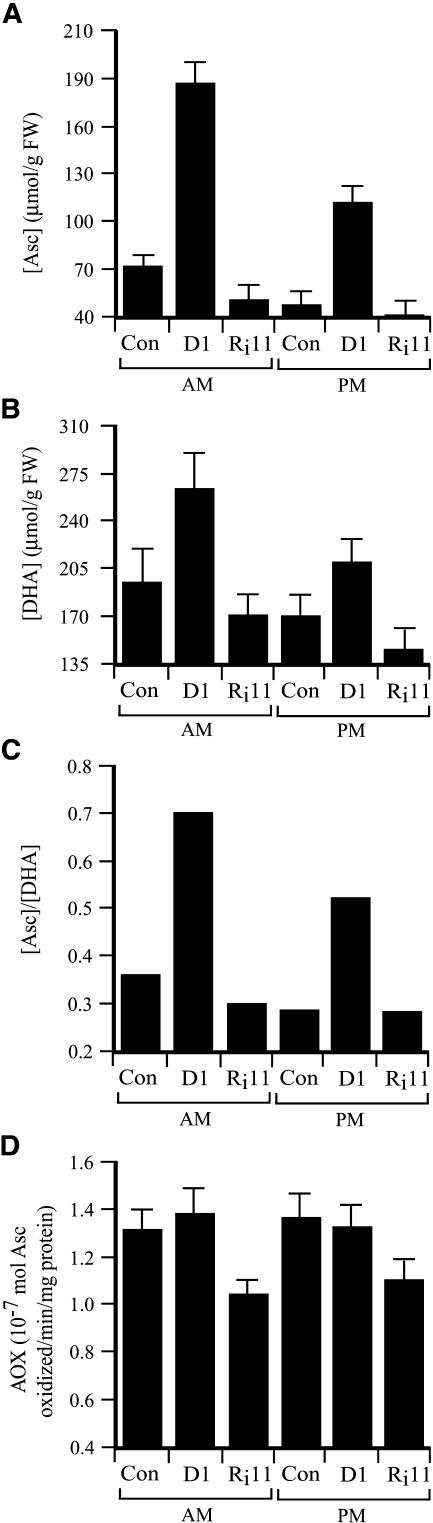
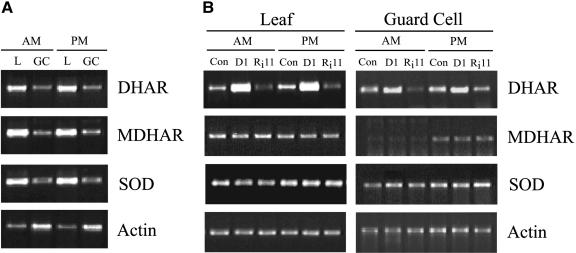
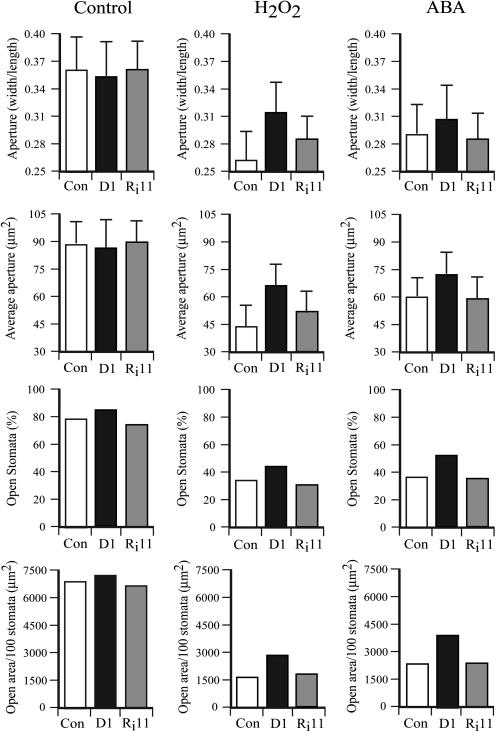
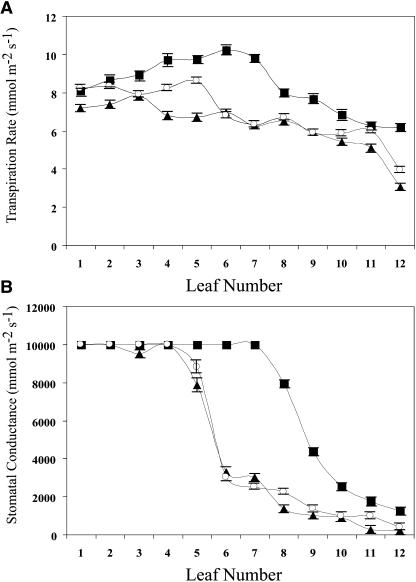
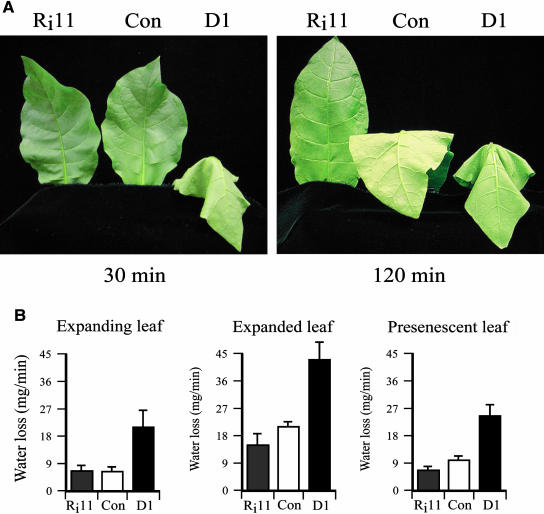
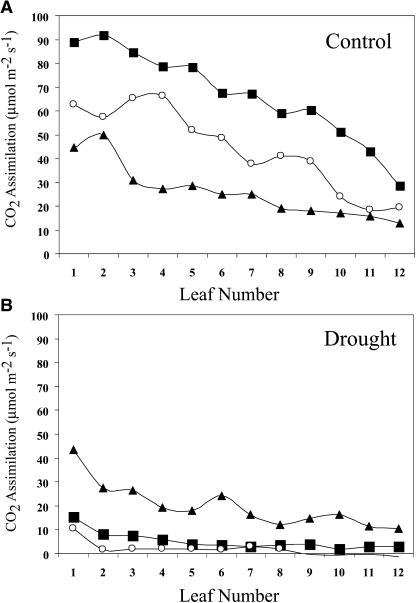
H(2)O(2) serves an important stress signaling function and promotes stomatal closure, whereas ascorbic acid (Asc) is the major antioxidant that scavenges H(2)O(2). Dehydroascorbate reductase (DHAR) catalyzes the reduction of dehydroascorbate (oxidized ascorbate) to Asc and thus contributes to the regulation of the Asc redox state. In this study, we observed that the level of H(2)O(2) and the Asc redox state in guard cells and whole leaves are diurnally regulated such that the former increases during the afternoon, whereas the latter decreases. Plants with an increased guard cell Asc redox state were generated by increasing DHAR expression, and these exhibited a reduction in the level of guard cell H(2)O(2). In addition, a higher percentage of open stomata, an increase in total open stomatal area, increased stomatal conductance, and increased transpiration were observed. Guard cells with an increase in Asc redox state were less responsive to H(2)O(2) or abscisic acid signaling, and the plants exhibited greater water loss under drought conditions, whereas suppressing DHAR expression conferred increased drought tolerance. Our analyses suggest that DHAR serves to maintain a basal level of Asc recycling in guard cells that is insufficient to scavenge the high rate of H(2)O(2) produced in the afternoon, thus resulting in stomatal closure.
Figures
References
-
- Aebi, H. (1984). Catalase in vitro. Methods Enzymol. 105, 121–126. - PubMed
-
- Asada, K. (1999). The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant Physiol. Plant Mol. Biol. 50, 601–639. - PubMed
-
- Assmann, S.M. (1993). Signal transduction in guard cells. Annu. Rev. Cell Biol. 9, 345–375. - PubMed
-
- Assmann, S.M., and Wang, X.Q. (2001). From milliseconds to millions of years: Guard cells and environmental responses. Curr. Opin. Plant Biol. 4, 421–428. - PubMed
-
- Blatt, M.R. (2000). Cellular signaling and volume control in stomatal movements in plants. Annu. Rev. Cell Dev. Biol. 16, 221–241. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
